盐碱胁迫冬油菜的主导因素分析
2013-03-14许耀照孙万仓曾秀存李彩霞周喜旺
许耀照,孙万仓,曾秀存,,李彩霞,周喜旺
(1.河西学院,甘肃 张掖 734000; 2.甘肃农业大学,甘肃 兰州 730070; 3.天水市农业科学研究所,甘肃 天水 741001)
我国约有667万hm2盐碱地[1],土壤盐化与碱化往往相伴发生,长期以来人们将土壤可溶性盐分的增加笼统地称为“土壤盐碱化”[2]。土壤盐化以土壤盐度升高为主要特征,碱化以土壤pH升高为主要特征[3],土壤盐渍化已成为当今世界农业发展所面临的重要问题[4],在我国主要的农业生产区河西走廊盐渍土面积达179.8万hm2[5],且土壤偏碱[6],开发利用河西走廊的盐碱地可以扩大耕地面积,提高农作物总产量。
近10年来,我国食用植物油的进口量持续增加,总进口食用植物油占中国食用植物油供给的62.41%[7],因此,在耕地有限的情况下,如何增加食用植物油自给显得尤为重要。油菜(Brassicacampestris)属于十字花科(Brassicaceae)芸薹属(Brassica),作为世界第3大油料作物和我国第1大食用植物油料作物[8-10],种植范围非常广泛,是生产优质植物油及优质饲料蛋白的重要作物。油菜一般被认为是中度耐盐作物[10]。随着冬油菜种植北移的成功[11],冬油菜在甘肃河西地区已成为一种重要的经济作物。目前关于盐碱混合胁迫的研究已在小麦(Triticumaestivum)[12-13]、野生大豆(Glycinesoja)[14]、甜高粱(Sorghumdochna)[15]和水稻(Oryzasativa)[16-17]等作物方面有报道,但关于油菜耐盐碱的研究多集中于NaCl等单一盐胁迫后一些生理生化指标的研究[9-10,18],而关于复合盐碱对冬油菜胁迫的主导因素研究很少。为了充分利用河西走廊的盐碱地种植冬油菜,增加食用植物油的产量,本研究依据河西走廊土壤盐碱特性,将中性盐NaCl和Na2SO4及碱性盐NaHCO3和Na2CO3按不同摩尔比例混合,模拟20种盐度和pH值不同的盐碱条件,对冬油菜“陇油6号”(Brassicacampestriscv.Longyou No.6)的种子和幼苗进行盐碱混合胁迫处理,测定种子发芽率、根系活力和叶片细胞膜透性等6项胁变指标,旨在探讨盐碱混合条件对冬油菜“陇油6号”胁迫作用的主导因素,为河西走廊盐碱地种植经济作物冬油菜提供理论依据。
1 材料与方法
1.1供试材料 超强抗寒冬油菜品种“陇油6号”由甘肃农业大学遗传育种教研室提供。试验于2010年3月到5月在河西学院农业与生物技术学院遗传学实验室中进行。
1.2试验设计 盐碱溶液的配制参照姚帅男等[19]的方法并加以改进,试验设计为4种盐分组成[A因素,以碱性盐所占摩尔比例由小到大分为4组。NaCl、Na2SO4、Na2CO3和NaHCO3的依次盐分组成摩尔比为A1(1∶9:9∶1)、A2(1∶1∶1∶1)、A3(9∶1∶1∶9)和A4(1∶1∶9∶9)]和5种盐浓度,10、20、30、40和50 mmol·L-1的双因素处理,总计20种盐度和pH值不同的盐碱组合。各处理液的pH值、盐度、Na+、Cl-、SO42-、HCO3-和CO32-的浓度根据盐处理液的实际盐分组成得出(表1)。
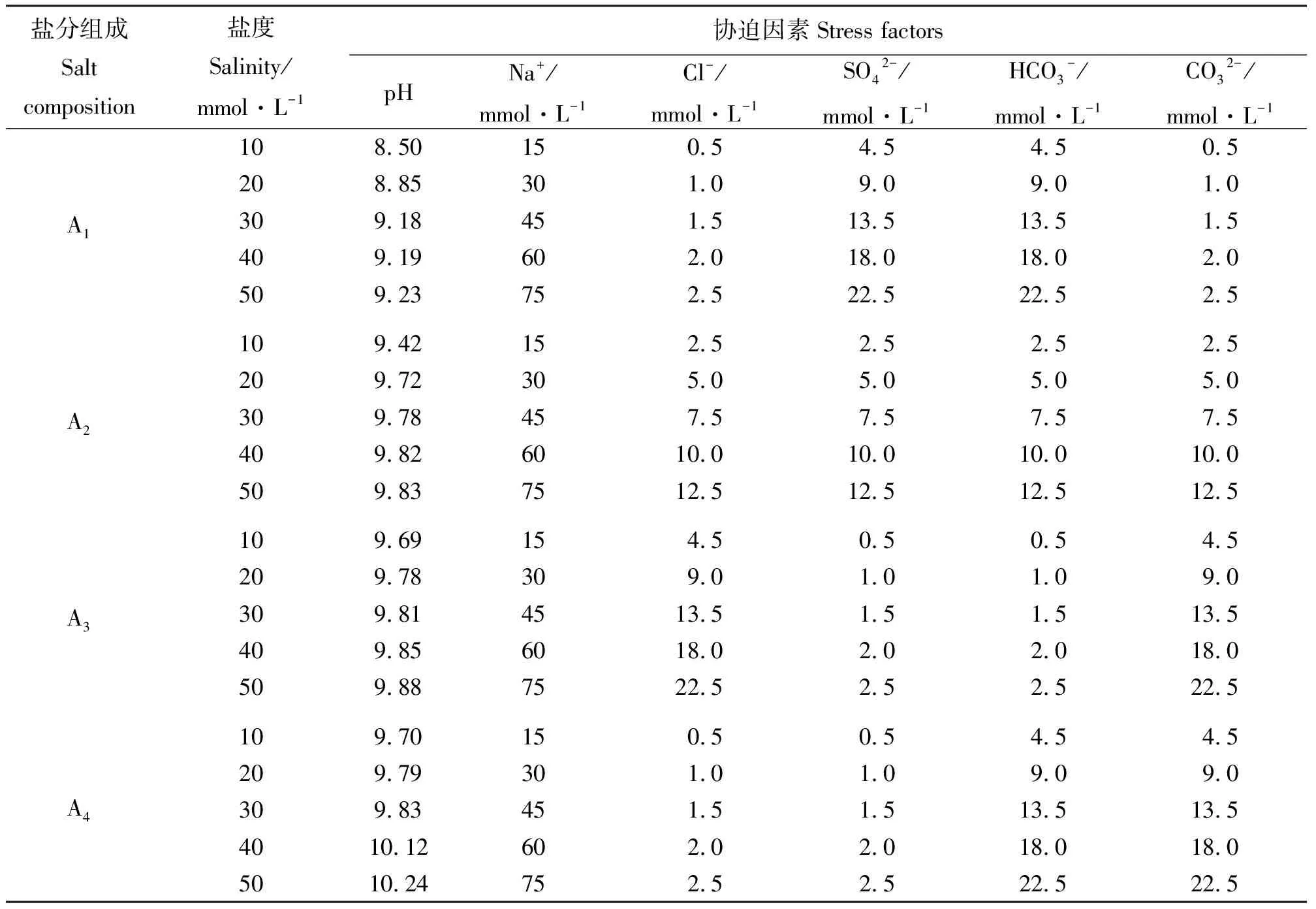
表1 不同盐处理液各胁迫因素的浓度Table 1 Density of stress factors for various salt treatments solution
1.3材料培养及测定项目 种子萌发:2010年3月6日挑选颗粒饱满、大小一致的“陇油6号”种子,用 0.1 % HgCl2消毒10 min,蒸馏水冲洗4次,用滤纸吸干种子外附水分,将种子置于不同盐碱混合处理溶液中浸种6 h,分别置于发芽床上,每个培养皿(Φ=12 cm)中放50粒种子,加入不同盐碱混合浓度的培养液10 mL,每个处理重复3次,用蒸馏水处理做对照(CK),然后将培养皿置于智能光照培养箱GZH-268B中,在25±1℃条件下避光培养。培养期间,每天定时观察,用称重法补充所蒸发的水分,每个培养皿补充一致,从处理的第2天开始,每天定时记录种子的发芽情况,连续统计7 d后,计算种子发芽率。
幼苗培养:2010年3月12日将消毒避光萌发的“陇油6号”种子,待胚根露出后播种至装有蛭石的花盆(Φ=15 cm)中,待两叶一心时,挑选萌发整齐,生长良好的幼苗,每盆5株继续培养。苗龄28 d时,选取长势均匀的63盆,随机分成21组,进行盐碱混合胁迫处理,每组3次重复,处理于18:00-20:00进行。以相应混合盐碱浓度溶液处理,每盆300 mL处理液分3次灌透,以保证油菜幼苗根系完全处于盐碱胁迫中,以蒸馏水处理为对照(CK),胁迫处理7 d后进行相关指标测定。取幼苗生长部位一致的第2片叶测定细胞质膜透性[20],用硫代巴比妥酸(TBA)法测叶片丙二醛含量[20],用磺基水杨酸法测定叶片脯氨酸含量[20],用考马斯亮蓝G-250法测定叶片可溶性蛋白含量[20],取幼苗根系生长部位一致的主根用氯化三苯基四氮唑(TTC)法测定根系活力[20]。
1.4数据分析 用CO32-、pH值、盐度和Cl-4因素代表所有的胁迫因素进行多元线性回归统计分析。以CO32-、pH值、盐度和Cl-为自变量,并设X1=CO32-、X2=Cl-、X3=pH、X4=盐度,X=b1X1+b2X2+b3X3+b4X4,b1~b4为回归系数;用发芽率、细胞膜透性、根系活力、丙二醛、脯氨酸和可溶性蛋白含量6项胁变指标为因变量Y,按公式Y=a+X对6项胁变指标进行多元回归分析,计算各自变量的标准回归系数b1′~b4′,以标准回归系数绝对值的大小直接反映胁迫因素对胁变指标的影响程度,并计算全相关系数的平方(R2)以评价各回归方程的回归效果。
数据用Microsoft Excel 2003软件进行处理和绘图,用DPS v 7.05数据处理软件进行相关性分析,试验数据为3次重复的平均值并进行Duncan氏新复极差法统计分析。
2 结果与分析
2.1不同盐分组成和不同盐浓度处理对胁变指标的影响 当盐处理液的浓度由10 mmol·L-1递增到50 mmol·L-1时,不同盐分组成的处理液A1~A4均降低了“陇油6号”种子发芽率、幼苗根系活力和叶片可溶性蛋白含量,增加了幼苗叶片细胞膜透性、丙二醛和脯氨酸的含量。在同一盐处理浓度下不同盐分组成的处理液之间各胁变指标的变化达显著性差异(P<0.05)(表2)。
随着盐分组成由A1到A4,“陇油6号”种子发芽率、幼苗根系活力和叶片可溶性蛋白含量大体降低,幼苗叶片细胞膜透性和脯氨酸含量大体增大,不同处理间差异显著(P<0.05)。当盐分组成为A1时,随着盐处理液浓度由10 mmol·L-1递增到50 mmol·L-1时,“陇油6号”种子发芽率由99.33%降低到96.00%、幼苗根系活力由120.76 μgTTF·g-1·h-1降低到94.56 μgTTF·g-1·h-1,叶片可溶性蛋白含量由8.93 mg·g-1降低到8.22 mg·g-1;幼苗叶片细胞膜透性由0.74%增加到0.79%、丙二醛含量由1.40 μmol·g-1增加到2.12 μmol·g-1和脯氨酸含量由21.73 μg·g-1增加到35.81 μg·g-1,不同处理间差异显著(P<0.05)(表2)。
当盐分组成为A4的50 mmol·L-1时,“陇油6号”种子的发芽率为22.67%、幼苗根系活力为85.58 μgTTF·g-1·h-1、叶片可溶性蛋白含量为7.25 mg·g-1、幼苗叶片细胞膜透性为0.84%、丙二醛含量为2.90 μmol·g-1和脯氨酸含量为41.75 μg·g-1,分别是盐分组成为A1的50 mmol·L-1时的23.61%、90.50%、88.20%、106.33%、136.79%和116.59%,这表明碱性盐摩尔比例增加明显影响各胁变指标的变化(表2)。
2.2协迫因素与协变指标间的相关性分析 pH值和CO32-与发芽率、细胞膜透性、丙二醛、根系活力、脯氨酸以及可溶性蛋白6项协变指标的相关系数均达到极显著水平(P<0.01)。从总体来看,CO32-最高,其次是pH值;在6项协变指标中,只有根系活力和脯氨酸含量分别与pH值和盐度的相关系数最高,其余各因素的最大相关因素均为CO32-,可见碱性盐对协变指标具有决定性作用。在各胁迫因素中,HCO3-除与脯氨酸含量具有显著相关性外(P<0.05),与其它各指标的相关系数都很低,而SO42-与各协变指标均无显著相关性,因此HCO3-和SO42-的致胁变作用可以忽略;Cl-除与发芽率无显著相关(P>0.05)外,与丙二醛具有极显著相关(P<0.01),与叶片细胞膜透性、脯氨酸含量、可溶性蛋白含量和根系活力协变指标呈显著相关(P<0.05),所以Cl-的胁迫作用不可忽略。本试验中,盐度和Na+完全正相关(r=1),盐度和Na+除与根系活力相关性较低外,与发芽率具有显著相关(P<0.05),与叶片细胞膜透性、丙二醛、脯氨酸和可溶性蛋白含量均达到极显著相关(P<0.01),盐度和Na+与各胁变指标的相关系数也一致,所以Na+的胁迫作用完全可由盐度代表(表3)。
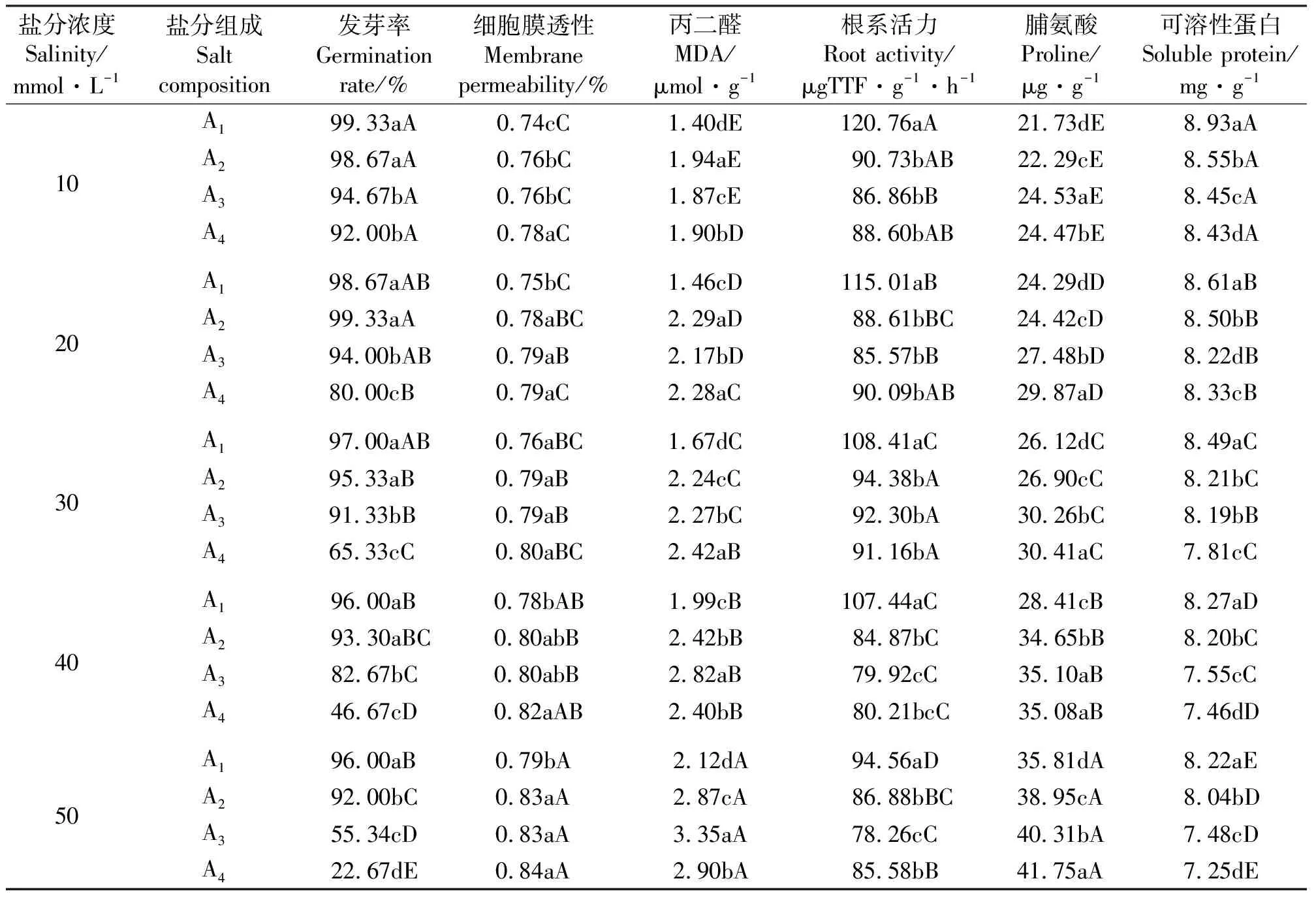
表2 不同盐分组合的处理液和不同浓度的盐处理液对胁变指标的影响Table 2 Effect of strain index on salt treatment with concentration and different concentrations of salt treatment
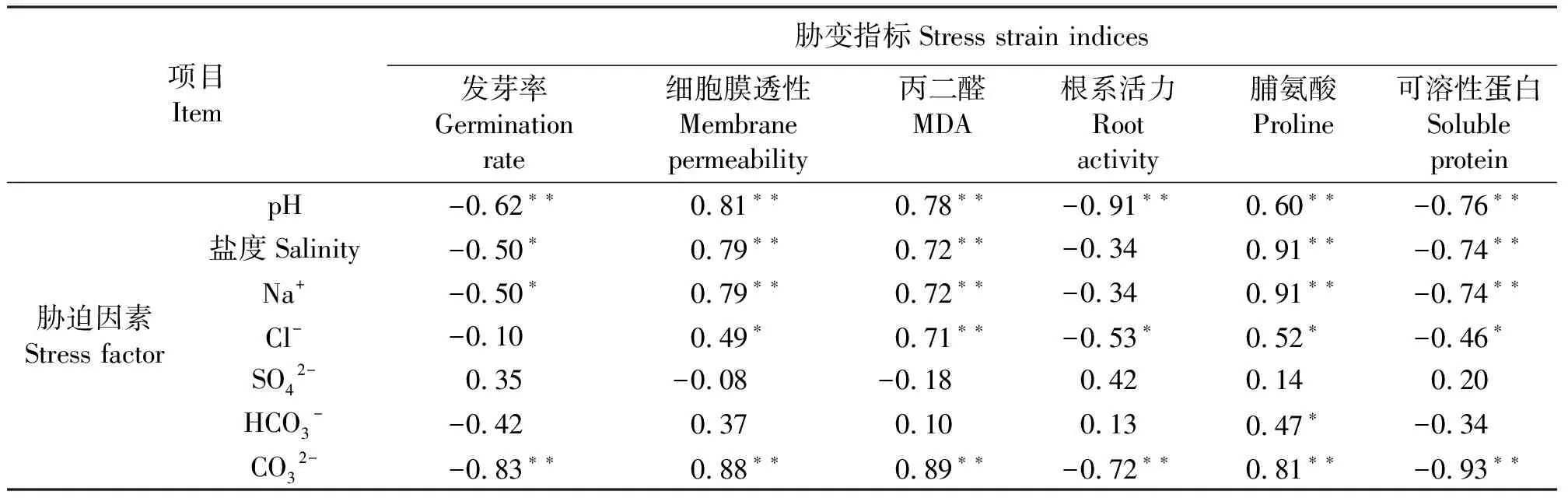
表3 胁迫作用因素与胁变指标间的相关性分析Table 3 Correlation coefficients between stress factors and strain indices
2.3胁迫因素与6项胁变指标间的多元回归分析 “陇油6号”种子发芽率与CO32-、Cl-、pH和盐度4因素间回归方程的全相关系数的平方R2为0.944 5,这表明4因素与种子发芽率间具有高度线性相关性。其它胁变指标的R2在0.873 3~0.959 4,分析结果均可证明,用CO32-、pH值、盐度和Cl-即可有效代表复合盐碱胁迫作用的各种因素。比较各胁迫因素的标准回归系数可知,对种子发芽率协变指标来说,各胁迫因素的重要性依次为 CO32->Cl->pH值>盐度,其中CO32-为决定性的主导因素,而盐度的作用较小,若去掉盐度进行三元回归分析得R2=0.942 3与四元回归效果基本一致。油菜叶片细胞膜透性四元回归R2为0.959 4,盐度为主要因素,而Cl-为最次要因素,所以将其去掉后进行三元回归分析得R2=0.936 6,与四元回归的效果基本一致,这证明了Cl-对细胞膜透性的影响较小。丙二醛含量四元回归R2为0.913 8;CO32-、Cl-、pH值和盐度4胁迫因素对MDA含量有同等重要的作用。根系活力与4胁迫因素R2为0.873 3,这表明4胁迫因素与根系活力间具有一定的线性相关性,比较各因素的标准回归系数可知,对根系活力胁变指标来说,各因素的重要性依次为pH值>Cl->CO32->盐度,其中pH值为决定性的主导因素,而盐度的作用较小,若去掉盐度进行三元回归分析得R2=0.871 7,与四元回归效果基本一致。脯氨酸含量四元回归的R2为0.937 9,盐度为脯氨酸含量的主要胁迫因素,其次是CO32-,而Cl-和pH值的作用较小,若去掉Cl-或pH值进行三元回归分析,其回归分析R2分别为0.936 9和0.937 2,这进一步证明了两者对脯氨酸含量影响较小。可溶性蛋白含量与4因素间具有高度线性相关性,各胁迫因素对可溶性蛋白含量的重要性依次为CO32-、盐度、Cl-和pH值,其中CO32-是决定性主导因素,而pH值的作用很小,去掉pH值后再进行三元回归分析的结果为R2=0.934 4,与四元回归分析的结果基本一致。由胁迫因素与胁变指标间的多元回归分析可得出,影响胁变指标的主要胁迫因素是CO32-、pH值和盐度(表4)。
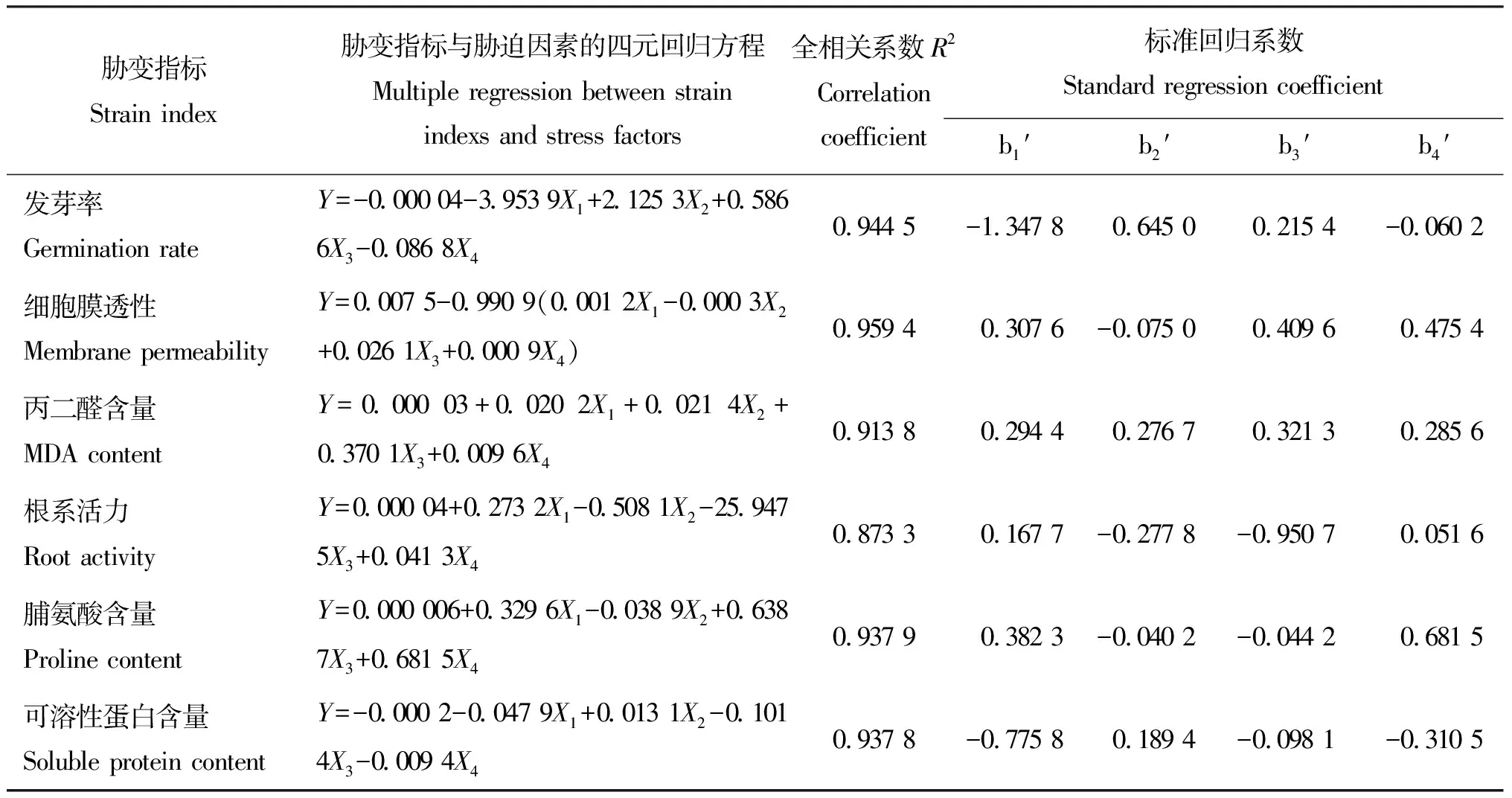
表4 胁变指标与胁迫因素间的多元回归分析Table 4 Multiple regression analysis between strain indexs and stress factors
3 讨论与结论
本试验依据甘肃河西地区盐碱地的组成特点[6]以及白菜型冬油菜的耐受能力,将NaCl、NaHCO3、Na2SO4和Na2CO3按不同摩尔比混合,模拟出20种低盐度、高pH盐碱生态条件,其盐度覆盖从10~50 mmol·L-1,pH值从8.50~10.24。一般中性盐NaCl的主要胁迫因子Na+可造成植物细胞水势下降的渗透效应以及以Na+为主的离子效应[8],而碱性盐Na2CO3的胁迫作用则需在前者的基础上加上高pH[9-10,12]。分析20种盐碱混合条件中的胁迫因素发现,pH值和CO32-与6项协变指标均达到极显著相关(P<0.01),盐度与Na+完全正相关,两者与各胁变指标的相关系数也一致,所以Na+完全可由盐度代替。Cl-与各胁变指标间具有极高的相关性,而HCO3-和SO42-与各胁变指标间的相关性很低,其致胁变作用可以忽略。试验结果表明,高盐和高碱的共同胁迫作用远大于单纯的高盐或高碱胁迫,表明盐度和碱度间具有明显的协同效应,应将碱性盐胁迫称为碱胁迫,中性盐胁迫称为盐胁迫,在生产中应将盐度和pH值作为评价土壤盐碱化程度的共同指标。
本研究表明,可用 CO32-、pH值、盐度以及Cl-4个胁迫因素代表盐碱混合对冬油菜的所有胁迫作用因素,这一结果与石德成等[2]的研究结果基本一致。通过对6项协变指标的回归分析发现,不同胁迫因素对6项协变指标的决定程度有所不同。发芽率和可溶性蛋白含量的主要胁迫因素均为CO32-,细胞膜透性和脯氨酸含量的主要胁迫因素均为盐度,而Cl-的胁变作用很小;根系活力与4个胁迫因素间回归分析发现pH值为决定性的主要胁迫因素,而盐度的作用较小;4个胁迫因素对丙二醛含量具有同等重要的胁变作用。在生产中应将pH值、盐度和CO32-作为冬油菜抗盐碱能力的测定指标。关于不同胁迫因素对不同胁变指标的影响程度的差异,可能与植物对胁迫响应的生理机制及胁变形成的生理过程有关,还需进一步试验研究。
盐度、pH值、CO32-和Cl-可代表盐碱混合条件下所有胁迫作用因素。pH值、盐度和CO32-是盐碱混合条件对冬油菜“陇油6号”胁迫的主导因素,共同影响着盐碱地冬油菜“陇油6号”的生长,同时Cl-的胁迫作用也是不可忽略的。
[1] 王玉祥,陈爱萍,张博.不同苜蓿品种耐盐性初探[J].草业科学,2010,27(3):102-106.
[2] 石德成,盛艳敏,赵可夫.复杂盐碱条件对向日葵胁迫作用主导因素的试验确定[J].作物学报,2002,28(4):461-467.
[3] 石德成,盛艳敏,赵可夫.复杂盐碱生态条件的人工模拟及其对羊草生长的影响[J].草业学报,1998,7(1):36-41.
[4] 吴凤萍,韩清芳,贾志宽.4个白花苜蓿品系种子萌发期耐盐性研究[J].草业科学,2008,25(8):57-62.
[5] 杨自辉,王继和,纪永福,等.河西走廊盐碱地治理模式研究[J].土壤通报,2005,36(4):479-482.
[6] 陈叶,罗光宏.甘肃河西地区盐碱地野生药用植物资源[J].干旱地区农业研究,2004,22(3):161-163.
[7] 陈允正,杨校生,格日乐图,等.中国主要油料作物及食用植物油产业变动特征[J].山西农业科学,2011,39(3):197-201,209.
[8] Becker H C,Loptien H,Robbelen G.Breeding:An overview[A].In:Gomez-Campo C.Biology ofBrassicaCoenspecies[C].Amsterdam,Elsevier Science Publishers,1999:413-460.
[9] 李源龙.油菜耐盐性的基因型差异[D].杭州:浙江大学,2007.
[10] 郑青松,刘海燕,隆小华,等.盐胁迫对油菜幼苗离子吸收和分配的影响[J].中国油料作物学报,2010,32(1):65-70.
[11] 孙万仓,武军艳,方彦,等.北方旱寒区北移冬油菜生长发育特性[J].作物学报,2010,36(12):2124-2134.
[12] 郭伟,于立河.盐碱胁迫对小麦幼苗根系活力和苯丙氨酸解氨酶活性的影响[J].作物杂志,2012(1):31-34.
[13] 郭伟,王庆祥,于立河.盐碱混合胁迫对小麦幼苗阳离子吸收和分配的影响[J].麦类作物学报,2011,31(4):735-740.
[14] 罗晓,曹蕾,王明超,等.野生大豆盐碱胁迫响应基因GsZFP1的克隆及序列分析[J].东北农业大学学报,2012,43(4):20-26.
[15] 戴凌燕,张立军,阮燕晔,等.盐碱胁迫下不同品种甜高粱幼苗生理特性变化及耐性评价[J].干旱地区农业研究,2012,30(2):77-83.
[16] 齐春艳,梁正伟,杨福,等.水稻耐碱突变体ACR78在苏打盐碱胁迫下的离子吸收特性[J].华北农学报,2009,24(2):112-116.
[17] 王志春,杨福,齐春艳.盐碱胁迫对水稻花粉扫描特征和生活力的影响[J].应用与环境生物学报,2010,16(1):63-66.
[18] 孟庆俊,冯启言,周东来,等.盐碱对绿豆和油菜种子萌芽的胁迫效应[J].安徽农业科学,2008,36(2):430,587.
[19] 姚帅男,刘晓东,郭林,等.金山绣线菊在混合盐碱条件下胁迫因素与胁变指标的相关性研究[J].北方园艺,2010(11):87-89.
[20] 王学奎.植物生理生化试验原理和技术[M].第二版.北京:高等教育出版社,2006.
