间伐对侧柏人工林碳储量以及细根生物量和形态的影响1)
2013-03-05李瑞霞彭婷婷闵建刚关庆伟
李瑞霞 彭婷婷 刘 娜 王 东 闵建刚 关庆伟
(南京林业大学,南京,210037)
森林是陆地生态系统中最大的碳库,其碳储量占全球陆地碳库总储量的46.27%[1]。因此,森林在应对全球气候变化过程中有不可替代的作用[2],而人工林作为陆地森林生态系统中重要的组成部分,其碳汇功能越来越受到重视[3-4]。然而,影响森林生态系统碳储量的因素很多,包括:气候[5],经营措施[6],土地利用方式[7-8],林分类型等[9-10],其中间伐是森林经营的重要措施。近年来,国内外研究间伐对森林碳储量的影响取得重大进展,但多限于植被层、土壤层和凋落物碳库变化结果的研究[11-16],对其变化机制的研究不多见[17-18]。根系中尤其是细根具有巨大的吸收表面积,生理活性强,是树木吸收水分和养分的主要器官,同时由于细根生长和周转迅速,每年对土壤有机碳库的贡献率高达25% ~80%[19],对森林生态系统碳分配和养分循环起着十分重要的作用[20-22]。间伐改变土壤养分含量、温度和水分等资源[23],这些因素的改变均可能引起细根结构和功能的改变[24-26]。其中细根生物量的变化反映了林木生产力和碳的分配格局的变化,但细根形态特征与其功能密切相关[27],故单独研究细根生物量不能充分反映其功能的变化。为此,本研究以徐州市石灰岩地区侧柏(Platycladus orientalis (Linn)Franco)人工林为研究对象,研究间伐对其碳储量、细根生物量和形态的影响,以期为侧柏人工林碳汇功能和科学经营提供基础数据和理论参考。
1 研究地概况
研究区位于江苏省徐州市的丘陵山地(116°22' ~118°40'E,33°43' ~34°58'N),山地面积3 万hm2,平均海拔400 m 以下。气候属暖温带半湿润季风气候区,四季分明,光照充足,雨量适中,年均温14 ℃左右,年均降水量800 ~900 mm,全年无霜期200 ~220 d。土壤以石灰岩发育而成的淋溶褐土为主,土层薄,砾石含量高,土壤黏重,中性至弱碱性。林分类型以侧柏人工林为主,还有小叶朴(Celtis bungeana Bl.)、麻栎(Quercus acutissima Carr.)、黄连木(Pistacia chinensis Bunge)、青桐(Firmiana simplex)、苦楝(Melia azedarach L.)、刺槐(Robinia pseudoacacia L.)和女贞(Ligustrum lucidum Ait.)等,而林下灌木和草本的发育较差。
2 材料与方法
2.1 样地设置
在林分踏查和标准地调查基础上,选择立地条件较接近的55年生侧柏人工林3 种间伐样地,分别为未间伐(CK),1998年实施1 次间伐(强度达11%),在原来的基础上于2005年实施第2 次间伐(间伐强度达43%),每种处理3 次重复,共设置固定样地9 块,每块样地面积25 m×25 m,林分概况详见(表1)。

表1 样地基本情况
2.2 侧柏人工林生态系统碳储量计算
利用李朝等的侧柏各器官生物量模型,计算了侧柏人工林乔木层的碳储量[28]。林下灌草层和枯落物的含碳率则采用常见的0.45 碳转化率[29],碳储量是根据单位面积林分干物质重量(生物量)乘以其碳含量求得:植物体碳储量(t·hm-2)=植物体生物量(t·hm-2)×植物体含碳率(%);土壤有机碳储量(t·hm-2)=土壤有机碳质量分数(g·kg-1)×土壤密度(g·cm-3)×土层厚度(cm)×10-1。
2.3 细根生物量和形态测定
2011年10月在每块标准地内随机选取3 个点,参考以往根系土块的取样方法[30-32],在离树50 cm 处,用平板利铲在0 ~10 cm 和>10 ~20 cm 的土层中各取20 cm ×20 cm 的土块,共54 块(9 ×2 ×3)。在实验室里,将根系用去离子水清洗干净,然后按照Pregitzer 等[30]的方法对结构完整的根进行分级。对结构不完整根系的细根根序等级依据完整根系的1 ~5 级根的长度和直径的范围来划分[31-32]。分级结束后,应用Epson 数字化扫描仪对分级后的不同处理样地的各级根序细根进行扫描,并采用根系分析系统软件WinRHIZO (Pro2005c)进行直径和根长的分析。扫描结束后,用滤纸包好各级根序,在65 ℃下烘干(48 h)至恒质量,用电子天平(±0.000 1 g)称质量。
2.4 数据处理
采用Excel 2003 进行数据整理和表格制作。运用SPSS13.0 分析不同间伐下侧柏人工林碳储量与细根形态和生物量有无显著性变化(p <0.05)及其相关性,多重比较采用Duncan 法。
3 结果与分析
3.1 不同间伐次数侧柏人工林生态系统碳储量分布格局
由表2可知,间伐后侧柏人工林生态系统碳储量分配格局为:土壤层>乔木层>枯落物层>灌木层>草本层,这与未间伐侧柏人工林分配格局基本一致。不同间伐侧柏人工林系统碳储量的变化规律为:2 次间伐>1 次间伐>未间伐,其中2 次间伐系统的碳储量为76.36 t·hm-2,分别比1 次间伐和未间伐林分多3.06、7.82 t·hm-2,这可能由于间伐后林分基本恢复,所以间伐林分系统碳储量大于未间伐林分的。1 次间伐后乔木层碳储量所占比例为50.96%,明显高于其他2 处理,这进一步说明间伐后乔木层的碳储量增加。2 次间伐后土壤层碳储量所占比例为51.57%,明显大于1 次间伐,这可能由于2 次间伐后林分中灌木、草本和枯落物增多,向土壤中输入的碳增多而致。灌木和草本层碳储量和占系统碳储量的比例均随间伐次数的增加而增加:2次间伐>1 次间伐>未间伐,这是由于间伐后,林内的光照增强,林木个体的营养空间增大,有利于林下灌木和草本的生长。因此,间伐后灌草层碳储量比未间伐的显著增加。枯落物层碳储量大小依次为:2次间伐>未间伐>1 次间伐,与土壤层碳储量的变化规律较一致,说明2 次间伐后枯落物增加,进而向土壤中输入的碳增多。
3.2 不同间伐次数对侧柏人工林细根形态的影响
由表3可知,与未间伐相比,1 次间伐后表层土壤中3、5 级根平均直径变细,4 级根直径变粗;亚表层土壤中1 ~5 级细根平均直径均变粗。与未间伐相比,2 次间伐后表层土壤中1 ~4 级根直径变粗,5级根直径显著变细(p <0.05);亚表层土壤中,3 ~5级根平均直径变粗,1、2 级根平均直径变化不大。与1 次间伐相比,2 次间伐后表层土壤中1 ~5 级根和亚表层4 级根平均直径均变粗,但亚表层2、3 和5 级根平均直径均变细。

表2 不同间伐次数侧柏人工林碳储量
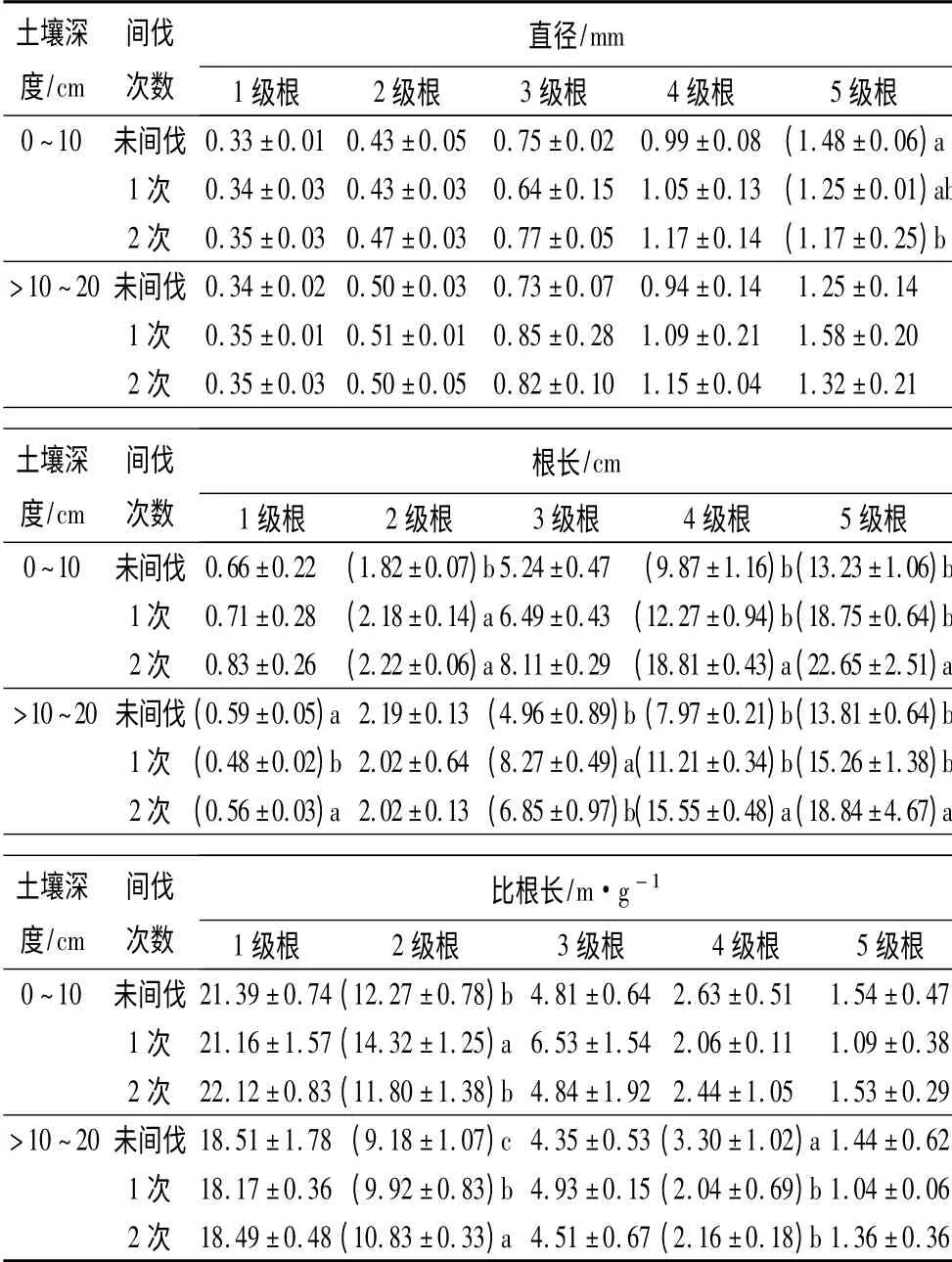
表3 间伐次数对细根形态的影响
由表3可知,与未间伐相比,1 次间伐后表层土壤1 ~5 级根平均根长均变长,其中2 级根的根长显著变长(p <0.05);亚表层土壤中1 级根的根长显著变短,而3 级根的根长显著增长(p <0.05)。与未间伐相比,2 次间伐后表层土壤中1 ~5 级根平均根长均变长,其中2、4 和5 级根平均根长显著增长(p <0.05);亚表层土壤中1 ~3 级根平均根长变化不显著(p >0.05),4、5 级根平均根长显著变长(p <0.05)。与1次间伐相比,2 次间伐表层土壤1 ~5 级根平均根长变长,其中4 级根平均根长显著变长(p <0.05);亚表层土壤中1、4 和5 级根平均根长变长,其中1 级根的平均根长显著增大(p <0.05)。
由表3可知,与未间伐相比,1 次间伐后,表层土壤中2 和3 级根的平均比根长增长,其中2 级根显著增长(p <0.05),4、5 级根的平均比根长变短,但无显著变化(p >0.05);亚表层土壤中1、4 和5 级根的平均比根长变短,但变化不明显(p >0.05),其中2 级根平均比根长显著增长(p <0.05),4 级根的显著变短(p <0.05)。与未间伐相比,2 次间伐后表层土壤中2 级根的平均比根长显著变短(p <0.05),其他根序间细根的平均比根长无显著变化,亚表层土壤中2 级根的平均比根长显著变长(p <0.05),4 级根的平均比根长显著变短(p <0.05)。与1 次间伐相比,2 次间伐后表层土壤2 级根的平均比根长显著变短(p <0.05);亚表层土壤中2 级根的平均比根长显著变长(p <0.05)。
3.3 不同间伐次数对侧柏人工林细根生物量的影响
由表4可知,与未间伐相比,表层土壤中2 次间伐显著增加了1 级根生物量(p <0.05),两种间伐显著降低了细根总生物量(p <0.05);亚表层土壤中2 次间伐显著增加了1 级和5 级根的生物量(p<0.05),两种间伐显著降低了4 级根的生物量(p<0.05)。从细根的垂直分布来看,各处理表层细根生物量均大于亚表层。以上结果说明,地上部分碳分配的增加而导致根系吸收能力的增加,植物体通过调整有限的碳源,使之分配到吸收功能最强的低级根,因此,间伐后低级根的生物量增加,而总生物量却显著减少。
3.4 不同间伐次数细根形态和生物量与生态系统碳储量的关系
合并0 ~20 cm 土层中细根形态和生物量指标并与生态系统各层碳储量做相关性分析(见表5),其中1 级根的直径与乔木层碳储量显著负相关;1 级根的长度与灌木层、枯落物层、土壤层和生态系统碳储量呈显著正相关;4 级根的比根长与土壤层碳储量呈正相关,5 级根的比根长与草本层碳储量呈显著负相关;1 级根的生物量与枯落物层碳储量呈极显著正相关,细根总生物量与乔木层碳储量呈显著负相关。以上研究表明,间伐后生态系统各层次碳储量的变化会显著影响1 级根的直径、根长和生物量,土壤层碳储量的变化显著影响4 级根的比根长,而草本层碳储量的变化显著影响5 级根的比根长。这一结果也充分证明了细根中低级根(1 ~3 级)的形态、结构决定其功能是吸收养分和水,寿命短、周转快;而高级根(4、5 级)的形态、结构决定其功能是运输和贮存养分,寿命长、周转慢[29-31]。林下枯落物量的多少及其分解快慢直接影响向土壤中输入养分从而间接影响了细根的形态和生物量,由此可见,适当的间伐不仅能改变碳储量的分配格局还能使整个生态系统碳储量增加。
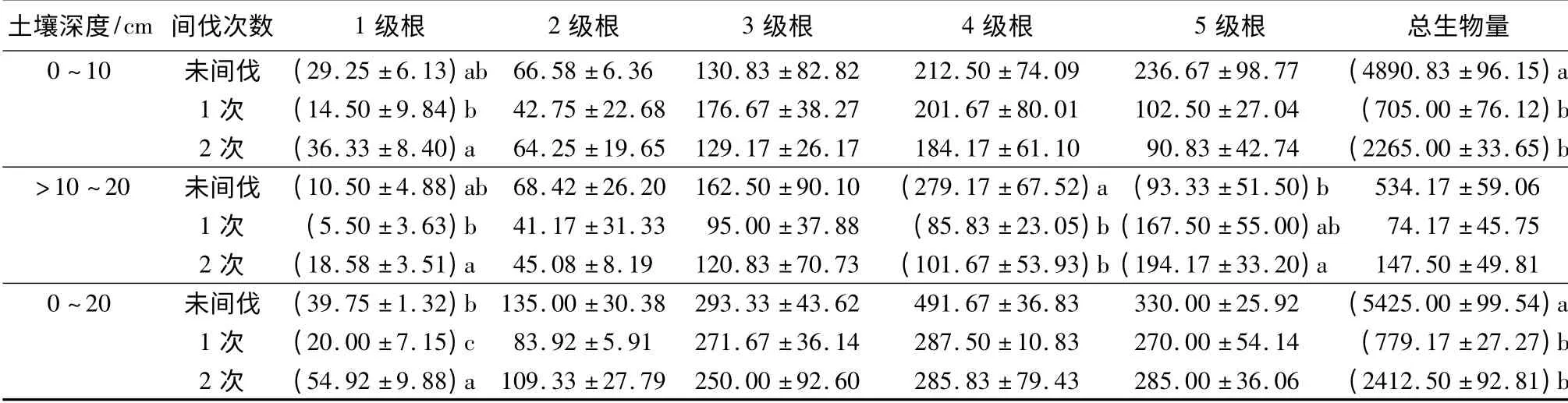
表4 间伐次数对细根生物量的影响 kg·hm -2
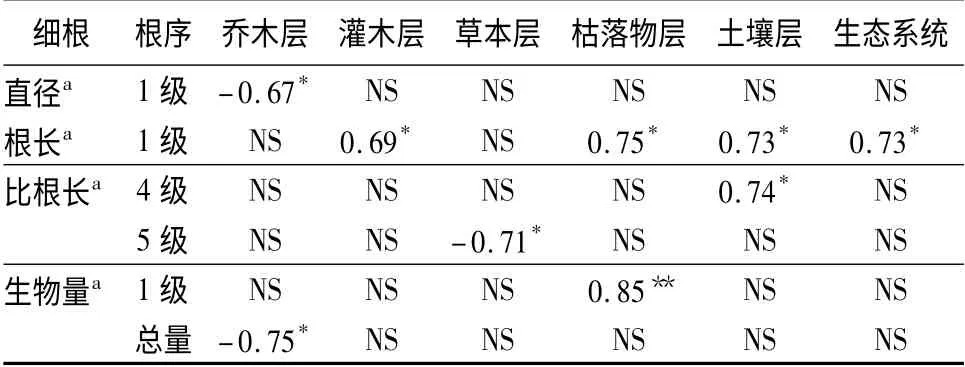
表5 不同林龄下细根形态、生物量与生态系统碳储量的关系
4 结论与讨论
侧柏人工林系统碳储量的变化规律为:2 次间伐>1 次间伐>未间伐,其中2 次间伐系统的碳储量为76.36 t·hm-2,分别比1 次间伐和未间伐林分多3.06 和7.82 t·hm-2。乔木层碳储量变化规律为:1 次间伐(37.35 t·hm-2)>2 次间伐(30.42 t·hm-2)>未间伐(28.13 t·hm-2)。土壤层碳储量变化规律为:2 次间伐(39.38 t·hm-2)>未间伐(36.09 t·hm-2)>1 次间伐(32.47 t·hm-2)。灌木层和草本层碳储量与系统碳储量的变化规律一致,枯落物层碳储量与土壤层碳储量的变化规律一致。可以预测,适度间伐强度可以增加系统碳储量。与未间伐相比,1 次间伐使表层土壤中2 级根的根长显著变长和亚表层土壤中1 级根的根长显著变短,表层和亚表层土壤中2 级根的比根长均显著变长。2次间伐后表层和亚表层土壤中4、5 级根的直径变粗、根长变长,表层土壤中2 级根的比根长变短,亚表层土壤中2 级根的比根长显著变长。与未间伐相比,2 次间伐显著增加了1 级根和亚表层5 级根生物量,而两种间伐显著减少了表层细根生物量,亚表层4 级根生物量和细根总生物量。生态系统各层次碳储量与1 级根的直径、根长和生物量显著相关,土壤层碳储量与4 级根的比根长显著正相关,而草本层碳储量与5 级根的比根长显著负相关。
本研究中,间伐后植被层碳储量增加,这与有关研究结果一致[15,33-35],但与方晰等[36]研究杉木人工林碳储量不同,这可能是因为间伐后林木株数的减少(林分密度下降),恢复时间或间伐强度的不同造成的。本研究表明,间伐显著减少了细根总生物量,这与相关的研究结果相同[23,37],而与另一些研究结果相反[14,24,38-39]。这与树种的萌发力和环境变化有关[24,37]。除此之外,此种差异可能与恢复时间的长短有关。本研究也表明,间伐对细根形态有显著影响,因细根的形态与其功能密切相关,如直径越小的细根的寿命越短[40-41]。由于根系形态具有高度的复杂性和可塑性,容易受土壤养分、水分以及温度的影响[42-44],随着时间的推移,不同间伐样地中的生物因子和非生物因子的差异也会影响细根结构和功能,进而对生态系统碳储量的空间分配格局产生影响。在本研究中,生态系统各层次碳储量与细根形态和生物量有一定的相关性,但因为本研究中所选侧柏人工林的样本数量较少,未来还应着重研究间伐后生态系统碳储量、细根形态和生物量的变化及其关系,为侧柏人工林的科学经营中确定适度的间伐强度或次数提供理论依据,充分发挥其生态效益和经济效益。
[1] IPCC. IPCC special report:land use,land-use change and forest[M]. Cambridge:Cambridge University Press,2000:4.
[2] Nowak D J,Daniel E C,Jack C S. Air pollution removal by urban trees and shrubs in the United States[J]. Urban Forestry & Urban Greening,2006,4:115 -123.
[3] 方精云,陈安平.中国森林植被碳库的动态变化及其意义[J].植物学报,2001,43(9):967 -973.
[4] Lal R. Forest soils and carbon sequestration[J]. Forest Ecology and Management,2005,220:242 -258.
[5] Cox P M,Betts R A,Jones C D,et al. Acceleration of global warming due to carbon-cycle feedbacks in a couple’s climate model[J]. Nature,2000,408:184 -187.
[6] 方晰,田大伦,项文化.不同经营方式对杉木林采伐迹地土壤C 储量的影响[J].中南林学院学报,2004,24(1):1 -5.
[7] Guo L B,Gifford R M. Soil carbon stocks and land use change:a meta analysis[J]. Global Change Biology,2002,8(4):345 -360.
[8] 吴建国,张小全,徐德应. 土地利用变化对土壤有机碳贮量的影响[J].应用生态学报,2004,15(4):593 -599.
[9] James W,Aydin T. Vegetation and soil respiration:correlation and control[J]. Biogeochemistry,2000,48:71 -90.
[10] 于贵瑞.全球变化与陆地生态系统碳循环与碳蓄积[M]. 北京:气象出版社,2003.
[11] Campbell J,Alberti G,Martin J,et al. Carbon dynamics of a ponderosa pine plantation following a thinning treatment in the northern Sierra Nevada[J]. Forest Ecology and Management,2009,257:453 -463.
[12] Chiang J M,McEwan R W,Yaussy D A,et al. The effects of prescribed fire and silvicultural thinning on the aboveground carbon stocks and net primary production of overstory trees in an oak-hickory ecosystem in southern Ohio[J]. Forest Ecology and Management,2008,255:1584 -1594.
[13] Hoover C,Stout S. The carbon consequences of thinning techniques:stand structure makes a difference[J]. Journal of Forestry,2007,105:266 -270.
[14] Argas R,Allen E B,Allen M F. Effects of vegetation thinning on above and belowground carbon in a seasonally dry tropical forest in Mexico[J]. Biotropica,2009,41:302 -311.
[15] 游伟斌,梁芳,贾忠奎,等. 抚育间伐对北京山区油松林乔木层碳储量的影响[J].北方园艺,2011(23):203 -206.
[16] Liang Fang,Ma Luyi,Jia Zhongkui,et al. Aboveground and root carbon stocks for Chinese arborvitae plantation following different silvicultural thinning[J]. Energy Procedia,2012,14:913-918.
[17] 王祖华,刘红梅,王晓杰,等. 经营措施对森林生态系统碳储量影响的研究进展[J]. 西北农林科技大学学报:自然科学版,2011,39(1):83 -88.
[18] Jandl R,Lindner M,Vesterdal L,et al. How strongly can forest management influence soil carbon sequestration?[J]. Geoderma,2007,137:253 -268.
[19] Usman S,Singh S P,Rawat Y S,et al. Fine root decomposition and nitrogen mineralization patterns in Quercus leucotrichophora and Pinus roxburghii forest in central Himalaya[J]. Forest Ecology and Management,2000,131:191 -199.
[20] 张小全,吴可红,Murach D.树木细根生产与周转研究方法评述[J].生态学报,2000,20(5):875 -883.
[21] Chen Xiaomei,Bledsoe C S. Contrasting seasonal patterns of fine root production for blue oaks (Quercus douglasii)and annual grasses in California oak woodland[J]. Plant and Soil,2002,240:263 -274.
[22] Kyotaro N,Tadashi S,Takeo M,et al. Estimation of the fine root biomass in a Japanese cedar (Cryptomeria japonica)plantation using minirhizotrons[J]. J For Res,2004,9(3):261 -264.
[23] Sword M A. Seasonal development of loblolly pine lateral roots in response to stand density and fertilization[J]. Plant and Soil,1998,200:21 -25.
[24] Tian Dalun,Peng Yuanying,Yan Wende,et al. Effects of thinning and litter fall removal on fine root production and soil organic carbon content in Masson pine plantations[J]. Pedosphere,2010,20(4):486 -493.
[25] López B C,Sabate S,Gracia C A. Thinning effects on carbon allocation to fine roots in a Quercus ilex forest[J]. Tree Physiology,2003,23:1217 -1224.
[26] Sword M A,Kuehler E A,Tang Zhenmin. Seasonal fine root carbohydrate relations of plantation Loblolly pine after thinning[J].Journal of Sustainable Forestry,2000,10(3/4):295 -305.
[27] Pregitzer K S,Jaskowski M J,Burton A J,et al. Variation in sugar maple root respiration with root diameter and soil depth[J]. Tree Physiology,1998,118:665 -670.
[28] 李朝. 侧柏人工林生物量研究[D]. 南京:南京林业大学,2010:6.
[29] Eggleston H S,Buendia L,Miwa K,et al. IPCC guidelines for national greenhouse gas inventories,prepared by the national greenhouse gas inventories programme[M]. Hayama:IPCC/IGES,2006.
[30] Pregitzer K S,Deforest J L,Burton A J,et al. Fine root architecture of nine North American trees[J]. Ecological Monographs,2002,72:293 -309.
[31] Guo Dali,Xia Mengxue,Wei xing,et al. Anatomical traits associated with absorption and mycorrhizal colonization are linked to root branch order in twenty-three Chinese temperate tree species[J]. New Phytol,2008,180(3):673 -683.
[32] 贾淑霞,赵妍丽,丁国泉,等. 落叶松和水曲柳不同根序细根形态结构、组织氮浓度与根呼吸的关系[J].植物学报,2010,45(2):174 -181.
[33] Sullivan B,Kolb T E,Hart S C,et al. Thinning reduces soil carbon dioxide but not methane flux from southwestern USA ponderosa pine forests[J]. Forest Ecology and Management,2008,255:4047 -4055.
[34] Hoover C,Stout S. The carbon consequences of thinning techniques:stand structure makes a difference[J]. Journal of Forestry,2007,105:266 -270.
[35] Finkral A J,Evans A M. Effects of a thinning treatment on carbon stocks in a northern Arizona ponderosa pine forest[J]. Forest Ecology and Management,2008,255:2743 -2750.
[36] 方晰,田大伦,项文化.间伐对杉木人工林生态系统碳贮量及其空间分配格局的影响[J]. 中南林业科技大学学报,2010,30(11):47 -53.
[37] Noguchi K,Han Q M,Araki M G,et al. Fine-root dynamics in a young hinoki cypress (Chamaecyparis obtusa)stand for 3 years following thinning[J]. Jouenal of Forest Research,2010,16(4):284 -291.
[38] Ryu S R,Concilio A,Chen J Q,et al. Prescribed burning and mechanical thinning effects on belowground conditions and soil respiration in a mixed-conifer forest,California[J]. Forest Ecology and Management,2009,257:1324 -1332.
[39] Hwang J,Son Y,Kim C,et al. Fine root dynamics in thinned and limed pitch pine and Japanese larch plantations[J]. J Plant Nutr,2007,30:1821 -1839.
[40] Wells C E,Eissenstat D M. Marked differences in survivorship among apple roots of different diameters[J]. Ecology,2001,82:882 -892.
[41] Matamala R,Gonzalez-Meler M A,Jastrow J D,et al. Impacts of fine root turnover on forest NPP and soil C sequestration potential[J]. Science,2003,302:1385 -1387.
[42] 康冰,刘世荣,蔡道雄,等.马尾松人工林林分密度对林下植被及土壤性质的影响[J].应用生态学报,2009,20(10):2323 -2331.
[43] Burton A J,Pregitzer K S,Hendrick R L. Relationships between fine root dynamics and nitrogen availability in Michigan northern hardwood forests[J]. Oecologia,2000,125(3):389 -399.
[44] Pregizer K S,King J S,Burton A J,et al. Research review:responses of tree fine roots to temperature[J]. New Phytol,2000,147:105 -115.
