中国刺槐次生种源对环境适应性比较分析1)
2013-03-05王进茂谷俊涛
赵 妙 张 军 王进茂 谷俊涛
(华北电力大学,保定,071001) (河北农业大学)
刺槐(Robinia pseudoacacia L.)是豆科双子叶植物,原产美国东部,17 世纪传入欧洲,19 世纪从欧洲传入亚洲[1]。刺槐在中国属于外来树种,经100 多年的种植几乎遍布于全国各地,面积在1 000 万hm2以上,已演化为我国的一个乡土树种。已有学者对我国刺槐进行了广泛研究,在刺槐的培育、养护和开发利用方面积累了丰富资料[2-6]。经多年发展,刺槐在我国已形成了次生种源。顾万春等[7]研究了我国9 个刺槐次生种源的遗传差异,首次证明我国刺槐次生种源经济性状存在真实遗传差异,并依据生产力和适应性评价,选择出3 个适应当地立地条件的优良次生种源。梁玉堂等[8]测试了我国9 个刺槐次生种源的抗热性生理指标,证明我国不同生态区的次生种源在高温适应性方面存在着差异。研究刺槐次生种源的分布和差异可以综合评价我国刺槐资源,揭示刺槐群体遗传多样性,为合理利用刺槐资源提供依据和材料。笔者从2003年开始搜集我国不同地区的19 个刺槐种源,并追踪其种源生长状况,对于刺槐生长性状的种源差异、种源与环境互作、种源稳定性及地理变异模式,以及我国刺槐群体的遗传结构和遗传多样性水平[9-11]的研究表明,刺槐种源间形成了明显的地理变异模式[12]。本文进一步研究19个刺槐种源在种植地的竞争适应变化,比较种源间竞争能力,以确定在种植地有发展潜力的种源。
1 材料与方法
1.1 种子收集
广泛搜集中国不同地区刺槐种子,包括吉林、辽宁、内蒙古、甘肃、山西、陕西、河北、河南、天津、山东、湖北、安徽、云南等省份,共收集次生群体刺槐种子19 份。
1.2 试验安排
2004年春季,在河北农业大学教学基地对19个刺槐次生种源群体进行育苗。2005年春天进行刺槐苗的移栽,在河北农业大学教学基地建立刺槐群体试验林,试验安排为:土壤质地为壤土,3月29日造林,供试的19 个刺槐群体在田间采用随机区组排列,小区面积92 m2,每个小区10 株,设3 次重复,每个种源初始数量为30 棵,行距0.8 m,株距0.6 m,4月5日平茬。
1.3 数据处理
自2005年起,每年秋天在刺槐落叶后,统计本年度刺槐存活的个体数,测定存活个体的树高(地面到植株最高处的距离)和胸径(地面1.3 m 植株的直径),分析各年度刺槐的生长和形态特征。采用Excel 软件和SPSS 统计软件对形态学性状变异进行分析。
19 个刺槐种源5 a 间各年度生长的存活率、树高和胸径等性状的显著性差异分析,选用方差分析或非参数检验Nemenyi 法进行种源间差异的多重比较(a=0.05)。
利用模糊数学中的隶属函数分析法,对19 个刺槐种源5 a 间各年度生长的存活率、树高和胸径等多个性状进行综合分析,进而做出每个群体的综合评价。
确定各性状的隶属函数值:
Xu=(X-Xmin)/(Xmax-Xmin)。
式中:X 为各个种源的某一性状的测定值;Xmax为所有种源中该测定值的最大值;Xmin为该性状中的最小值。若呈负相关,则Xu=1 -(X -Xmin)/(Xmax-Xmin)。
将每个种源各性状的隶属函数值进行累加,计算其平均值。平均值越大,说明种源的生长状况越好,即该种源对环境的适应及竞争能力越强,反之则弱。
2 结果与分析
2.1 刺槐不同群体生长状况
自2005年至2009年秋天,连续5 a 测定刺槐各个种源的生长状况。每年各个种源的存活个体数、存活率及多重比较Nemenyi 检验结果列于表1。
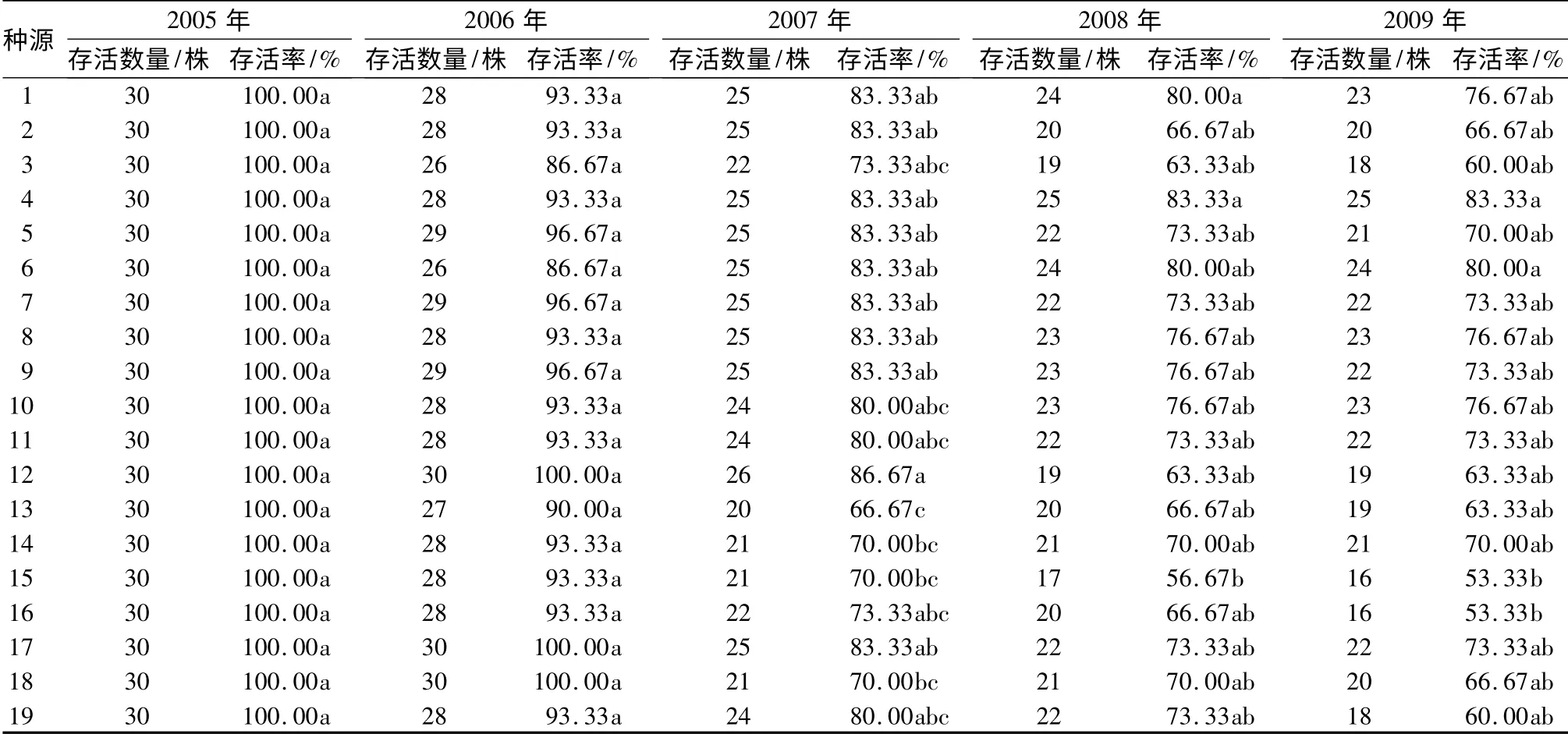
表1 19 个刺槐种源各年度的存活数量及存活率
从2005年春季平茬至秋天落叶,经1 a 的生长,19 个刺槐种源的存活率均为100%。在2006年,19个刺槐种源中种源12、种源17 和种源18 的存活率为100%,其余16 个种源都出现了死亡个体,但19个种源的存活率没有显著性差异。在2007年,19 个刺槐种源中部分种源的存活率出现了显著性差异,其中:种源12 的存活率最高,为86.67%;种源13 的存活率最低,为66.67%。2008年和2009年,19 个种源的存活率都呈现下降趋势,部分种源的存活率达到显著性差异,但种源间的存活率差异性表现为不稳定变化,如种源12 和种源13 的存活率在2007年表现为显著性差异,而在随后的2008年和2009年,两种源的存活率又表现为差异不显著。在2009年,种源4 的存活率最高,为83.33%;种源15 和种源16 的存活率最低,为53.33%。
19 个刺槐种源经过3 个年度的生长,存活率出现显著性差异。由于每个种源都有各自的遗传结构组成,而且个体间存在差异,在种植密度大的情况下,随着个体的生长,对环境资源要求加大,个体之间出现对环境资源的竞争,竞争力弱的个体被淘汰。19 个种源间存活率差异性出现的不稳定变化,可能是因为各个种源的个体尚处于幼年阶段,未形成绝对的生长优势,随着竞争的进一步加大,种源间的存活率仍在变化,即经过5 个生长年度仍不能确定19个刺槐种源存活率的固定差异性。
2.2 刺槐不同种源生长变异
统计19 个刺槐种源第5年生长的树高与胸径,并计算19 个种源树高与胸径的均值、最大最小值、标准差以及变异系数,将19 个种源的树高和胸径均值分别进行非参数多重比较Nemenyi 检验和方差分析,比较结果见表2。
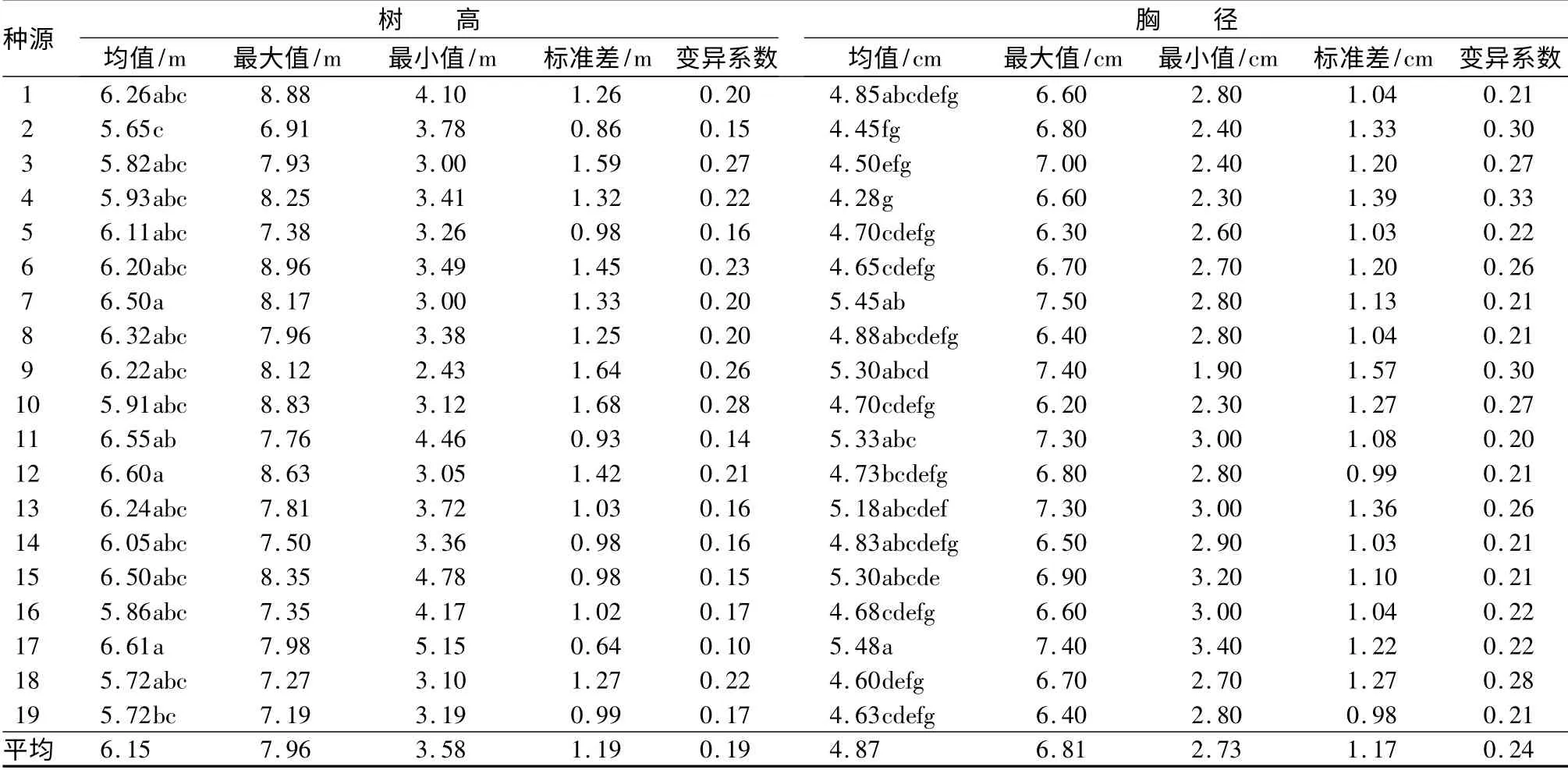
表2 2009年度19 个种源树高与胸径基本数据
经过5 个年度的生长,19 个刺槐种源的树高平均值为6.15 m,树高平均值范围为5.65(种源2)~6.61 m(种源17),种源间的树高平均值差异性达显著性水平,每一个种源内个体间的树高差异性更是达显著性水平。19 个刺槐种源的胸径平均值为4.87 cm,胸径平均值范围为4.28(种源4)~5.48 cm(种源17),种源间的胸径平均值差异性达显著性水平,每一个种源内个体间的胸径差异性同样达显著性水平。
种源间和种源内树高和胸径的差异代表了刺槐群体间和个体间生长的差异。种源内个体间的生长差异一方面受自身遗传的影响,在环境资源紧张时还与自身的竞争能力有关。经过5 个年度的生长,种源17 的树高平均值和胸径平均值都最大,分别为6.61 m 和5.48 cm;种源2 的树高平均值最小,为5.65 m,种源4 的胸径平均值最小,为4.28 cm。但种源4 的存活率比种源17 高,因此,要比较种源间对环境适应性能力,还要综合考虑各个种源的形态生长性和存活率的差异。
2.3 刺槐种源综合评价
采用隶属函数值法对19 个刺槐种源进行综合评价,得出19 个刺槐种源的形态(树高、胸径)数据和存活率3 个方面的隶属函数值的综合排序位次,结果见表3。
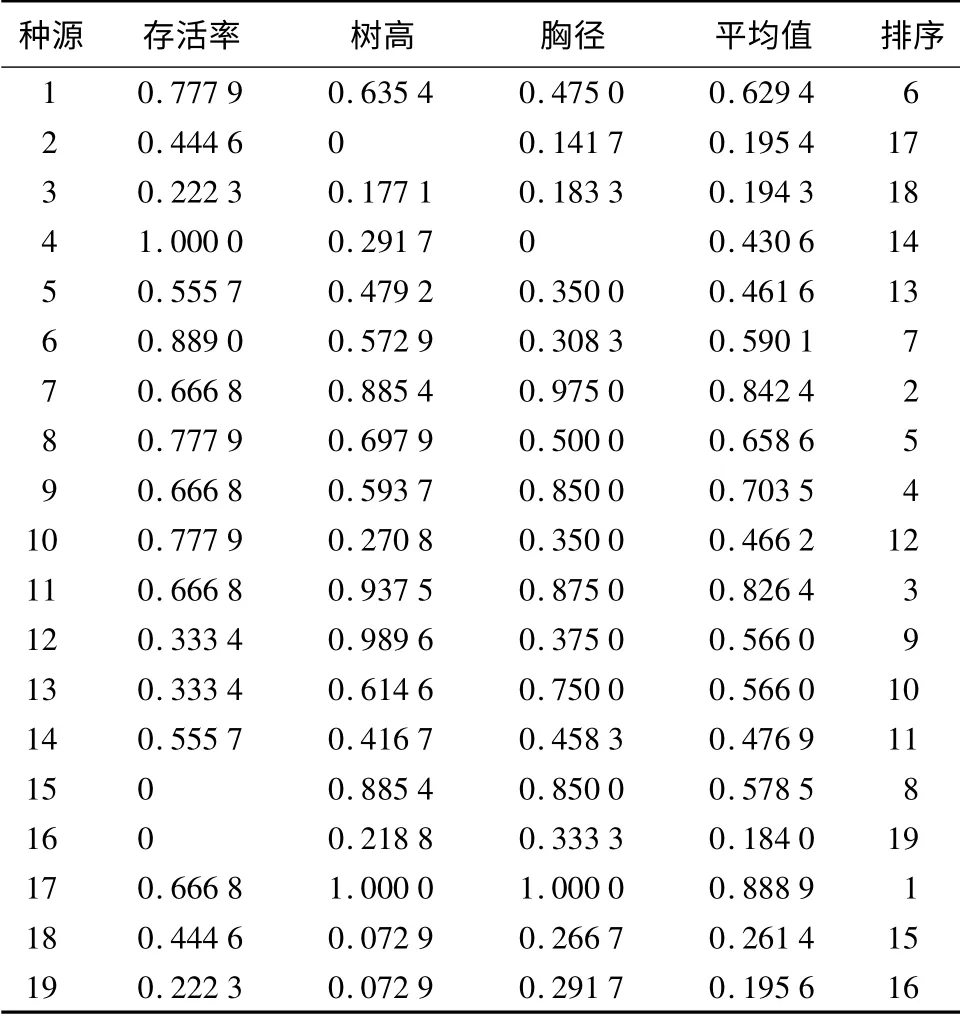
表3 19 个刺槐种源的生长状况的隶属函数值及综合评价
综合考虑各个种源的树高、胸径以及存活率,种源17 的隶属函数平均值最大,为0.888 9;种源16的隶属函数平均值最小,为0.184 0。种源17 的平均树高和胸径在19 个种源中均最大,存活率为73.33%,在19 个种源中也比较高,因而表现最为突出,说明该种源在本地适应程度最好;而种源16 在19 个种源中的存活率最小,为53.33%,平均树高和胸径虽然不是最小,但树高和胸径的平均值与最小树高和胸径的平均值无显著差别,说明种源16 在本地适应程度最差。
3 结论与讨论
生物通过适应环境而存在,植物群体形态特性是植物群体基因型适应当地环境的综合表现,分布范围广的物种,其生长特性和遗传组成往往呈现典型的地理变异模式。文献报道[13],不同产地的树种种源存在显著的遗传差异并多呈纬向变异模式,且低纬度的种源生长速度快于高纬度的种源。在中国,刺槐属外来树种,种植有100 多年历史,已演变为乡土树种,且形成了次生种源[7-8]。王玉等[12]对种植在石家庄高邑、邢台沙河和保定市3 个试验点的我国19 个刺槐次生种源4年生的试验林进行了调查,研究了其生长性状方面的种源差异、种源与环境互作、种源稳定性及地理变异模式,发现刺槐树高性状不存在显著的种源差异,而胸径在种源之间差异显著,并运用AMMI 模型及Eberhart & Russell 进行了联合回归分析,表明刺槐种源间形成了明显的地理变异模式。本文比较并分析了我国19 个刺槐次生种源种植在河北省保定生态条件下5 个年度的存活率及形态生长特性,种源17 的隶属度平均值最大,种源16 的隶属度平均值最小,初步表明种源17在本地适应程度最好,而种源16 在本地适应程度最差。19 个刺槐次生种源在河北省生态条件经过5个年度的生长,未表现出明显的地理变异模式,可能是因为19 个刺槐次生种源生长年度较短,种源个体间仍有激烈竞争,自我稀疏选择还在进行的缘故,另外,从树高和胸径的标准差可以看出,当种源存活率基本稳定时再分析19 个刺槐次生种源的形态生长特性,其地理变异模式可能会更清晰。
物种长期适应不同自然环境条件可产生丰富的种内遗传变异,使得物种往往存在显著的种源间和种源内个体间的性状差异,形成适应一定生态环境的生态型。特定的生态型有一定的遗传结构,影响一个物种遗传结构的内部因素有突变、交配系统、遗传漂变、基因流动等,而外部因素如自然选择及人类影响等对物种群体的遗传结构也有显著影响[14-16]。逆境对植物群体遗传结构有选择作用,在研究盐胁迫对刺槐群体遗传结构的影响[17]中发现,盐胁迫下刺槐群体的SSR 标记遗传参数发生了改变,随着盐浓度的增加,刺槐群体中平均位点杂合体越来越多,说明盐胁迫对刺槐群体的遗传结构有定向选择作用。在利用等位酶标记对本文19 个刺槐群体的遗传结构和遗传多样性水平的研究中,通过与欧洲刺槐群体的比较,发现我国刺槐群体具有较高的遗传多样性,在等位酶水平未形成明显的地理变异规律或模式[9,18]。本研究中的19 个刺槐次生种源在河北保定生态条件下会有重新适应的变化,在高密度种植条件下的5 个试验年度中,随着个体的生长,种源内和种源间竞争激烈,竞争存在自疏现象,竞争能力强的个体存活下来,经过自然选择与淘汰,刺槐群体的遗传结构可能会发生改变。对比研究19 个刺槐次生种源遗传结构经过5 个年度的变化,有利于分析群体遗传结构的变化规律,发掘和利用适应特定环境的刺槐次生种源和种源内的优良个体。
[1] Keresztesi B.刺槐[M].王世绩,张敦伦,译.北京:中国科学技术出版社,1993.
[2] Boring L R,Swank W T. The role of black locust (Robinia pseudoacacia)in forest succession[J]. Journal of Ecology,1984,72(3):749 -766.
[3] 中国树木志编辑委员会.中国刺槐造林技术[M]. 北京:中国林业出版社,1978.
[4] 山东省林业研究所.刺槐[M].北京:农业出版社,1974.
[5] 彭鸿,陈晓荣,余仲东. 刺槐人工林培育实践的认知[J]. 水土保持学报,2003,17(5):11 -15.
[6] Xie Yan,Li Zhenyu,William P G,et al. Invasive species in China-an overview[J]. Biodiversity & Conservation,2001,10(8):1317 -1341.
[7] 顾万春,王全元,张英脱,等.刺槐次生种源遗传差异及其选择评价[J].林业科学研究,1990,3(1):70 -75.
[8] 梁玉堂,丰震,龙庄如,等.刺槐次生种源抗热的研究[J].山东农业大学学报,1993,24(5):126 -133.
[9] Gu Juntao,Yang Minsheng,Wang Jinmao,et al. Genetic diversity analysis of black locust (Robinia pseudoacacia L.)distributed in China based on allozyme markers approach[J]. Frontiers of Agriculture in China,2010,4(3):366 -374.
[10] 孙芳,杨敏生,张军,等.刺槐不同居群遗传多样性的ISSR 分析[J].植物遗传资源学报,2009,10(1):91 -96.
[11] 韩宏伟,张世红,徐兴兴,等. 中国刺槐种源间抗寒性地理变异研究[J],河北农业大学学报,2008,32(2):57 -60.
[12] 王玉,谷俊涛,习洋,等. 中国刺槐次生种源生长变异研究[J],河北农业大学学报,2011,34(2):27 -33.
[13] 王秀花,陈柳英,马丽珍,等.7年生木荷生长和木材基本密度地理遗传变异及种源选择[J],林业科学研究,2011,24(3):307 -313.
[14] Loveless M D,Hamrick J L. Ecological determinants of genetic structure in plant population[J]. Annual Review of Ecology and Systematics,1984,15(1):65 -95.
[15] Hamrick J L,Godt M J W,Sherman-Broyles S L. Factors influencing levels of genetic diversity in woody plant species[J]. New Forests,1992,6(1/4):95 -124.
[16] Waples R S,Gaggiotti O. INVITED REVIEW:What is a population?An empirical evaluation of some genetic methods for identifying the number of gene pools and their degree of connectivity[J]. Moleclar Ecology,2006,15(6):1419 -1439.
[17] Gu Juntao,Liu Weina,Akindele Akinnagbe,et al. Effect of salt stress on genetic diversity of Robinia pseudoacacia seedlings[J].African Journal of Biotechnology,2012,11(8):1838 -1847.
[18] 杨敏生,Hertel H,Schneck V.欧洲刺槐种源群体遗传结构和多样性[J].生态学报,2004,24(12):2700 -2706.
