利用内含子切接点引物分析西藏青稞种质资源的遗传多样性
2013-02-27曾兴权王玉林徐齐君原红军尼玛扎西
曾兴权 王玉林 徐齐君 原红军 尼玛扎西
(1.西藏自治区青稞种质改良和牦牛繁育重点实验室,西藏拉萨850002;2.西藏自治区农牧科学院,西藏拉萨850002)
利用内含子切接点引物分析西藏青稞种质资源的遗传多样性
曾兴权1,2王玉林1,2徐齐君1,2原红军1,2尼玛扎西1,2
(1.西藏自治区青稞种质改良和牦牛繁育重点实验室,西藏拉萨850002;2.西藏自治区农牧科学院,西藏拉萨850002)
采用26条内含子切接点引物(ISJ),对50份来自藏区的青稞育成品种、50份西藏近缘野生大麦、44份西藏青稞地方品种,共144份供试材料的遗传多样性进行了分析。结果分析表明,筛选利用22对多态较好的引物或引物组合对实验材料进行扩增,不同引物或引物组合扩增条带数变幅为3~12。144份材料间GS值变化变幅为0.06~0.96之间;群体内的平均GS大小顺序依次为,西藏青稞地方品种>西藏近缘野生大麦>藏区青稞育成品种;遗传多样性分析顺序依次为西藏近缘野生大麦(1.5964)>藏区青稞育成品种(1.5841)>西藏青稞地方品种(1.1657)。对统计条带并建立[1,0]矩阵,聚类与主坐标分析表明,以144份材料的GS值0.40(L)为阈值,可将其划分为七类。研究结果表明,同一来源地区的青稞育成品种遗传基础较为狭窄,而不同地区间的藏区青稞育成品种遗传差异较大;在进行青稞超高产新品种选育时,应充分发掘西藏近缘野生大麦和利用不同地域来源的育成品种,加强不同地区间资源的交换,以利于增加群体材料的遗传多样性和新品种的培育。
青稞;种质资源;遗传多样性;内含子切接点引物
中国青藏高原是栽培大麦的发源地,也是品种多样性的重要地区,其中主要栽培的作物就是青稞。青稞(Hordeum vulgare L.var.nudum HK.f.2n=14)又称裸大麦,青藏高原在3 500年以前,青稞就作为西藏农牧民主要的粮食作物和重要的畜牧饲料,而如今青稞同时作为健康产品和酿造业创造价值;青稞在西藏种植面积最大(青稞种植面积为21万~22万hm2),分布最广(分布于全区28个农业县),品种类型最多[1]。
青藏高原生态类型繁多,不同区域降雨、温度、土壤类型、植被和紫外线辐射各有不同,这赋予青稞具有高原极端条件下广泛的适应性和多样性;同时,西藏农业历史悠久,最能适应高原生态环境的青稞种质资源极为丰富。从青藏高原及周边地区不同的地理区域,大量的育成品种、地方品种和野生青稞品种被收集起来,显示出丰富的形态和遗传变异、生物胁迫耐受[2-4]。随着人口增加、环境破坏、气候变化,人类对粮食作物需求增加。而这些青稞品种勿庸置疑是丰富宝贵的基因资源,为基因的挖掘和作物改良提供条件。遗传多样性是生物多样性的重要组成部分,是物种多样性的基础,在揭示物种或种群的进化史的同时,使我们发现更丰富的遗传资源,应用于农作物品种改良。
在已有的研究中,RFLP标记、RAPD(randomly amplified polymorphic DNA)标记、AFLP标记、SSR(Simple SequenceRepeats,SSR)标记、ISSR标记(Inter-Simple Sequence Repeat ISSR)等广泛地应用于植物遗传多样性分析。内含子在绝大多数植物中都存在,同时,外显子与内含子的结合区域高度保守,相对于外显子,通常内含子所受到的选择压力较弱,在不同物种甚至不同个体间,内含子碱基序列和长度变异较大[16]。由于(intron-splice junction primer,ISJ)可根据内含拼接位点设计正向引物和反向引物,而且引物间可以任意组合,已广泛地应用于动物、酵母、植物等遗传多样性研究中,均取得了很好的效果[17-19]。已有学者研究了高原地区青稞农艺性状[6]、贮藏蛋白[7-8,12]、RAPD[9]、SSR[2,10-11,13]、ISJ[14]、SRAP[15]。但是利用ISJ标记分析西藏青稞地方种质资源、西藏野生大麦群体、青藏区育成品种内以及这几个群体间的比较及关系研究报道较少。
本文利用ISJ标记技术对西藏种质资源不同类型群体的遗传多样进行研究,探讨这些资源群体内和群体间的遗传差异,明确西藏近缘野生大麦与西藏青稞地方品种、藏区青稞育成品种的关系,从资源角度对青稞种质资源材料进行综合评价,以促进丰富的青稞种质资源在西藏青稞的高产、超高产育种与遗传改良中的有效利用。
1 材料与方法
1.1 试验材料
实验共选取144份材料,其中由西藏自治区农牧科学院提供的藏区青稞育成品种50份、西藏青稞地方品种44份,由中国农业科学院作物所提供的西藏近缘野生大麦50份。材料具体名称见表1。
1.2 基因组DNA提取、引物设计、PCR扩增及产物检测、数据分析
引物采用Song等[17]设计的ISJ引物,结合利用曾兴权等[19]的设计的引物。基因组DNA提取、PCR反应体系、电泳、照相及数据分析均参照曾兴权等[19]的方法进行。
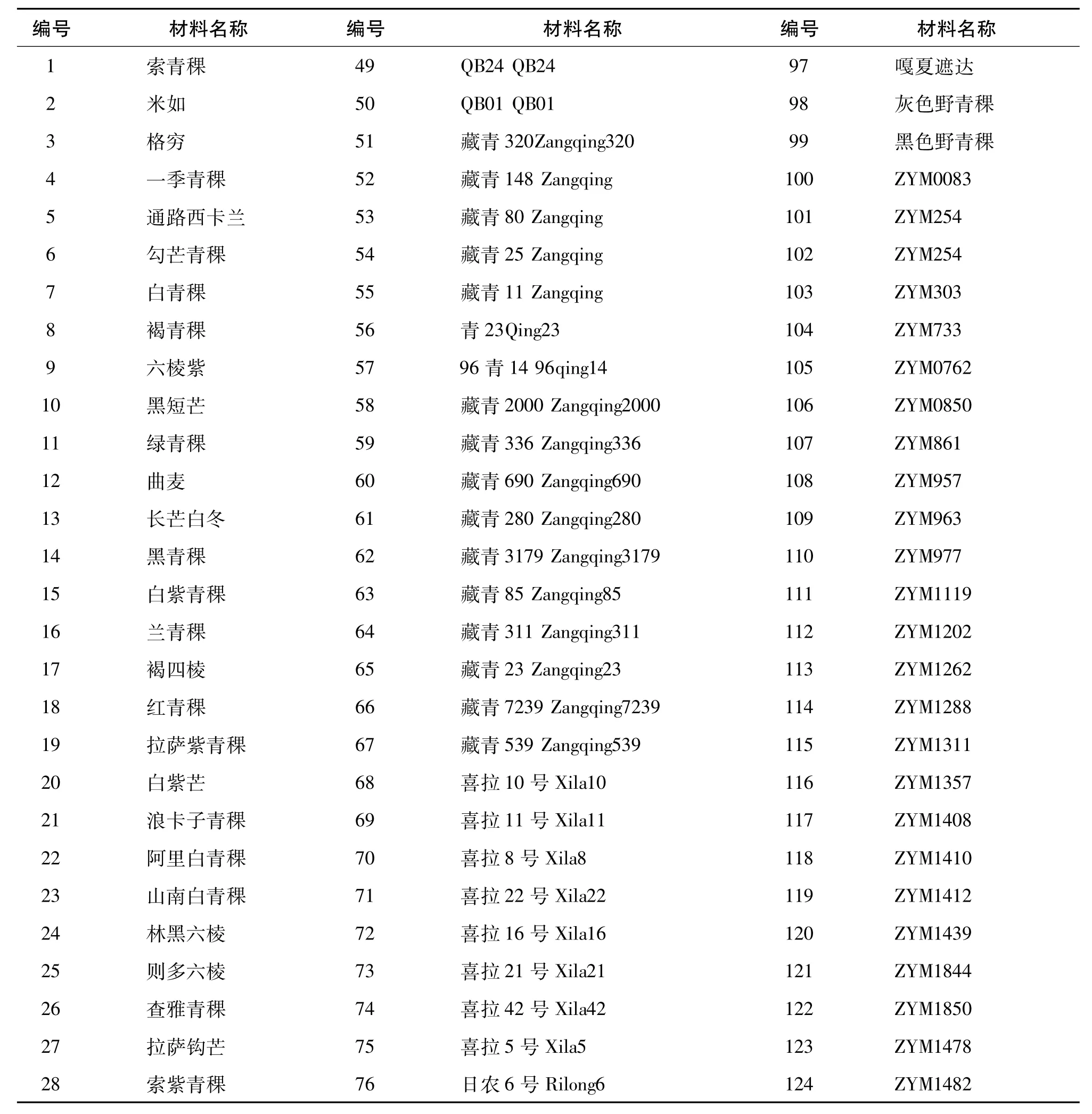
表1 144份供试材料
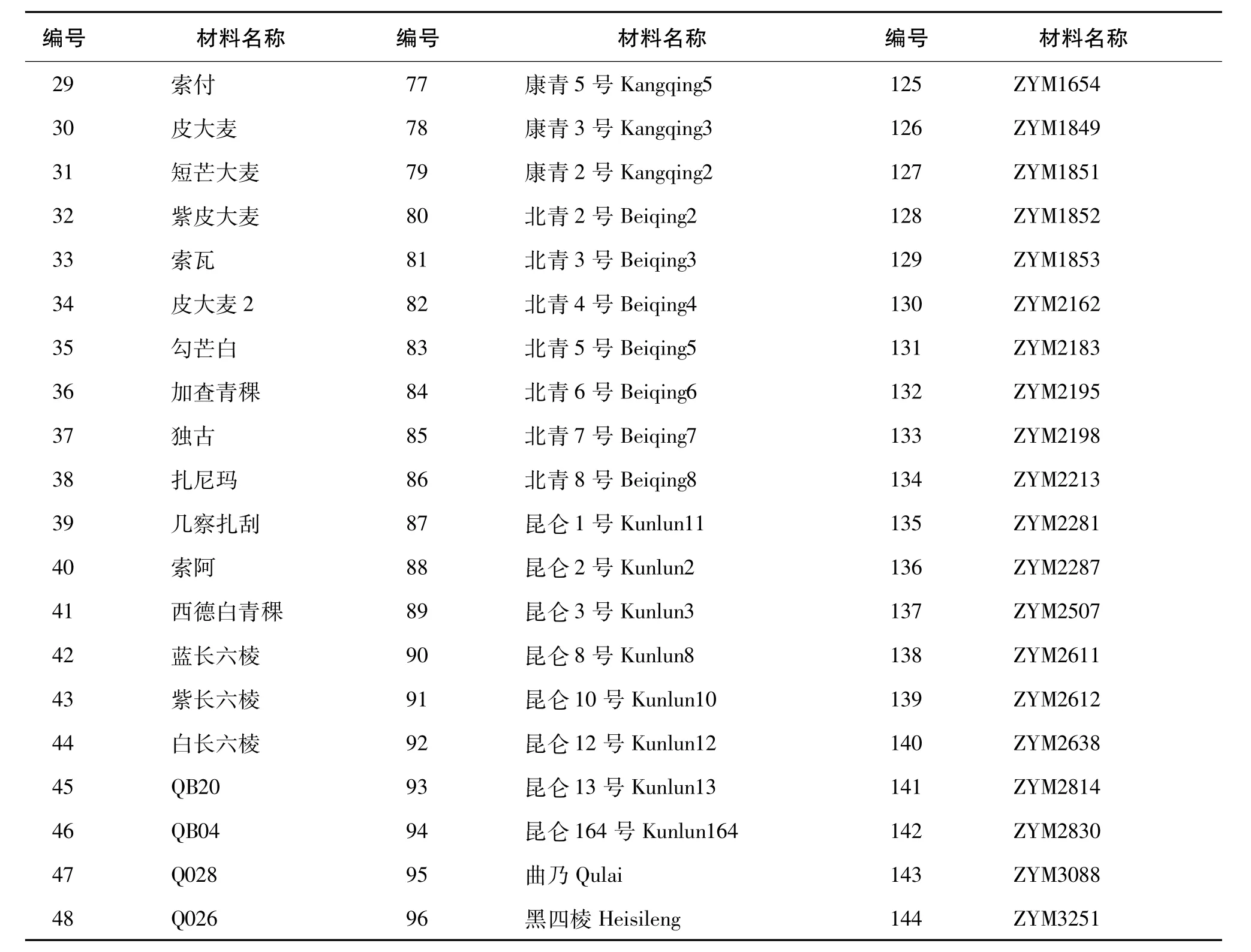
续表1

表2 内含子切接点引物序列
2 结果与分析
2.1 试验材料扩增产物的多态性
本研究利用26条ISJ引物对供试材料进行了多态性扩增,实验中共得到50个多态性引物或引物组合,占筛选引物的25%。选取其中22对多态较好的引物或引物组合对实验材料进行扩增。不同引物或引物组合扩增条带数变幅为3~12,引物A1检测出的等位变异最多,为12个等位变异,H11检测出的等位变异最少,为3个等位变异。在西藏近缘野生大麦、西藏青稞地方品种、藏区青稞育成品种平均分别为5.8、5.5、5.7条。在西藏近缘野生大麦中能扩增出127条稳定清晰的条带,多态性条带比例(PPB)为94.8%;在西藏青稞地方品种能扩增出稳定清晰的121条带,PPB为90.3%;在藏区青稞育成品种能扩增出125条稳定清晰的条带,PPB为93.3%;扩增结果见图1。
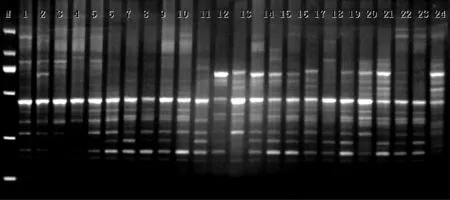
图1 西藏青稞种质资源H8引物部分扩增图
2.2 不同种质材料之间的遗传相似性比较
依据144份材料PCR扩增的结果构建[1,0]矩阵,计算供试材料间的Dice遗传相似系数(GS),实验结果表明,144份材料间GS值变化变幅为0.06~0.96之间。其中来自西藏近缘野生的灰色野青稞与黑色野青稞间的GS值最大,遗传相似程度最高。西藏青稞地方品种通路西卡兰和格穷与西藏青稞育成品种藏青690之间的GS值最小,遗传相似程度最低,因而遗传距离最远。144份材料间GS值的变幅为0.06~0.96,平均为0.540 2;其中西藏近缘野生大麦材料间GS值的变幅为0.13~0.93平均为0.539 9;西藏青稞地方品种间的GS值变幅为0.64~0.96,平均为0.732 2;藏区青稞育成品种材料间GS值的变幅为0.08~0.87,平均为0.536 5。以上结果表明,群体内的平均GS顺序依次为,西藏青稞地方品种>西藏近缘野生大麦>藏区青稞育成品种。这些说明了西藏青稞地方品种比西藏近缘野生大麦与藏区青稞育成品种具有更高的GS值,遗传相似程度较高,材料间遗传距离也相应较近;同时也表明藏区青稞育成品种和西藏近缘野生大麦群体内部遗传差异小于西藏青稞地方品种。
2.3 不同材料之间的遗传多样性比较及聚类分析
遗传多样性指数(H')分析结果表明,144份材料的H'为1.684 8。其中西藏青稞地方品种材料群体的H'为1.165 7,西藏近缘野生大麦材料群体的H'为1.596 4,藏区青稞育成品种材料群体的H'为1.584 1。这些结果表明,西藏近缘野生大麦材料和藏区青稞育成品种材料的遗传多样性大于西藏青稞地方品种。
聚类分析表明,校验得到的符合系数为0.905 6,表明构建的聚类图与获得的数据具有较好的符合度。聚类图分析结果表明,以144份材料的平均遗传相似系数0.40(L)为阈值,可将其划分为七类。进一步分析发现第I类有134份材料,其中青稞育成品种46份,西藏青稞地方品种38份,西藏野生大麦50份;第II类包括1份材料,为西藏青稞地方品种;第III类包括3份材料,全部为青稞育成品种;1份西藏青稞地方品种归属为第Ⅳ类;各2份西藏青稞地方品种分别归属为第Ⅴ、Ⅵ类;1份藏区青稞育成品种归属为Ⅶ。进一步从主坐标分析(图2)表明,所得结果与聚类分析完全一致。
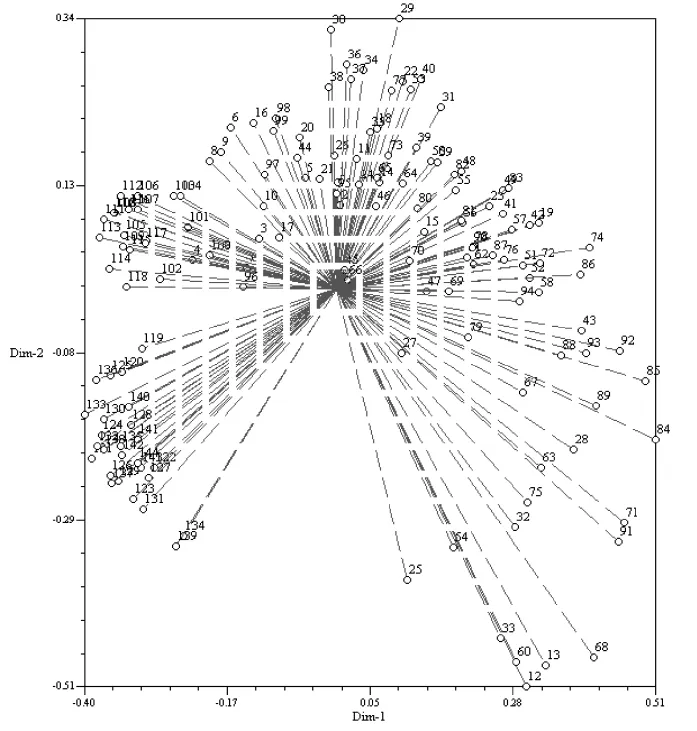
图2 144份材料主要坐标分析
3 讨论
遗传多样性研究是作物遗传育种的基础,对西藏青稞种质资源进行遗传多样性研究已成为重要课题之一。深入的研究藏区青稞育成品种、西藏青稞地方品种、西藏近缘野生大麦的遗传多样性,有利于发掘潜在的遗传优异基因资源。
吴昆仑[11]利用14对SSR引物研究认为青藏高原青稞遗传基础较广,蕴藏着多种不同的等位基因,具有丰富的遗传多样性,可为青稞重要性状遗传特性、基因资源挖掘和品种选育等方面的研究提供参考。曾兴权等[13]利用260对SSR引物对来自青藏高原的175份青稞种质资的遗传多样性进行了分析,结果表明同一来源地区的青稞育成品种遗传基础较为狭窄,不同地区间的青稞育成品种遗传差异较大;在西藏青稞新品种选育和遗传改良中,发掘和利用西藏野生大麦资源、西藏青稞地方品种资源的同时,更应该加强不同地区青稞育成品种资源的交换和配合使用,有利于增加群体材料的遗传多样性和新品种的培育。潘志芬等[12]利用SSR标记分析了64份青藏高原栽培青稞的遗传多样性,认为青藏高原栽培青稞具有丰富的遗传多样性;杨平等[15]利用SRAP分子标记技术,对25份来自四川高原的青稞育成品种进行了遗传多样性研究,结果表明,材料聚类与其来源地有明显的相关性;25份材料间的平均遗传距离较小,平均遗传多样性较低,遗传基础较为狭窄。张镝等[20]采用AFLP标记对西藏近缘野生大麦进行了遗传多样性分析,结果表明西藏近缘野生大麦具有丰富的遗传多样性。汪爱华等[21]采用RAPD和ISSR标记对110份青藏高原近缘野生大麦材料进行了遗传多样性分析,结果表明西藏地区近缘野生大麦具有较高的遗传多样性。
本研究认为,西藏青稞地方品种比西藏近缘野生大麦与藏区青稞育成品种具有更高的GS值,遗传相似程度较高,材料间遗传距离也相应较近;同时也表明藏区青稞育成品种和西藏近缘野生大麦群体内部遗传差异小于西藏青稞地方品种;西藏近缘野生大麦的平均遗传多样性高于藏区青稞育成品种及西藏青稞地方品种的平均遗传多样性。本文的结果进一步证实了以往的研究[13],同一来源地区的青稞育成品种遗传基础较为狭窄;不同地区间的青稞育成品种遗传差异较大;我们在研究这些材料的醇溶蛋白遗传多样性时,也发现了此规律,青藏高原栽培青稞具有丰富的遗传多样性。在进行青稞超高产新品种选育时,应充分发掘西藏近缘野生大麦和利用不同地域来源的育成品种,加强不同地区资源的交换和配合使用,有利于增加群体材料的遗传多样性和新品种的培育[10-11,13]。
[1]胡颂杰.西藏农业概论[M].四川科学技术出版社,1995,144-145
[2]潘7志芬,邹弈星,邓光兵,等.青藏高原栽培青稞SSR标记遗传多样性研究[J].中山大学学报(自然科学版),2007,46(2):81-87
[3]Huaxin Dai,Wunian Shan,Jing Zhao,et al.Difference in response to aluminum stress among Tibetan wild barley genotypes[J].Journal of Plant Nutrition and Soil Science,2011,174(6),952-960
[4]Wang,C.P.Pan ZF,Nima ZX,etal.Starch granuleassociated proteins of hull-less barley(Hordeum vulgare L.)from the Qinghai-Tibet Plateau in China[J].Journal of Sci.Food Agric.2011,91,616-624
[5]齐军仓,张国平.大麦醇溶蛋白与麦芽品质关系的研进展[J].麦类作物学报,2005,25(3):115-118
[6]孟凡磊,赵亚斌,强小林,等.不同地区大麦品种农艺性状比较与西藏青稞品种改良[J].麦类作物学报,2006,26(5):175-177
[7]周洪金,刘新春,刘仙俊,等.川藏高原地区青稞育成品种醇溶蛋白遗传多样性的比较分析[J].大麦与谷类科学,2007,(4)1-5
[8]刘新春,苟琳,杨平,等.青藏高原青稞品种醇溶蛋白的遗传多样性[J].植物遗传资源学报,2008,9(2):180-185
[9]洪棋斌,侯磊,罗小英,等.应用RAPD分析川西北高原青稞的遗传背景[J].中国农业科学,2001,34(2):133-138
[10]杨菁,迟德钊,吴昆仑,等.青海省栽培青稞SSR标记遗传多样性研究[J].安徽农业科学,2010,38(8):4307-4309
[11]吴昆仑.青稞种质资源的SSR标记遗传多样性分析[J].麦类作物学报,2011,31(6):1030-1034
[12]潘志芬,邹弈星,邓光兵,等.青藏高原青稞B组醇溶蛋白遗传多样性研究[J].应用与环境生物学报,2006,12(5):601-604
[13]曾兴权,王玉林,徐齐君,等.利用SSR引物分析西藏青稞种质资源的遗传多样性[J].麦类作物学报,2013,33(2):260-267
[14]孟凡磊,强小林,佘奎军,等.西藏主要农区青稞品种的遗传多样性分析[J].作物学报,2007(33):1910-1914
[15]杨平,刘仙俊,刘新春,等.利用SRAP标记研究四川高原青稞育成品种的遗传多样性[J].遗传,2008,30(1):115—122
[16]Hawkins,J.D.A survey on intron and exon length.Nucleic Acidsy Research.1998,16:9893-9905
[17]Song Wei-Ning,Langridge P.Identification and mapping polymorphism in cereals based on polymerase chain reaction.Theor Appl Genet.1991(82):209-216
[18]de Barros LM,Soden A,Henschke P A,Langridhe P.PCR differentiation of commercial yeast strains using intron splic site primers.Appl Environ Microbiol,1996,62:4514-4520
[19]Xingquan Zeng,Yajuan Wang,Weiyan Li,et al.Comparison of the genetic diversity between Triticum aestivum ssp.tibetanum Shao and Tibetan wheat landraces(Triticum aestivum L.)by using intron-splice junctionprimers[J].Genet Resour Crop Evol,2010,57:1141-1150
[20]张镝,丁毅.基于AFLP标记的中国西藏近缘野生大麦遗传多样性分析[J].遗传,2007,29(6):725-730
[21]汪爱华,丁毅.西藏近缘野生大麦RAPD和ISSR分子标记的遗传多样性[J].武汉大学学报(理学版),2007,53(6):723-730
Assessment of Genetic Diversity of Tibetan Hulless Barley Germ plasm(Hordeum vulgare L.)by ISJ Primers
ZENG Xing-quan1,2WANG Yu-lin1,2XU Qi-jun1,2YUAN Hong-jun1,2NIMazhaxi1,2
(1.Barley Improvement and Yak Breeding Key Laboratory of Tibet Autonomous Region,LhaSa 850002,China;2.Tibet Academy of Agriculture and Animal Husbandry Sciences,Lhasa 850002,China)
In order to assess the rich Tibetan Hulless barley germplasm in Tibet,we analyzed the genetic diversity of144 Hulless barley varieties,including 50 Tibetan bred barley materials,50 wild relatives of barley materials,and 44 Tibet highland barley landrace varieties by 26 introns splice junction(ISJ)primers.The results showed that there were 22 primerswith high polymorphism outof26 ISJ primers,bands ranged from 3 to 12 and the GS values ranged from 0.06 to 0.96 among the 144 materials.The average size of GS sequence in different population of hulless barleys was Tibet highland barley landrace varieties>wild relatives barley>Tibetan bred barley.The results of those analyses on the genetic diversity indicated thatwild relatives of barley varieties(1.5964)>Tibetan bred barley(1.5841)>Tibet highland barley landrace varieties(1.1657).Clustering and main coordinate analysis showed that when the threshold value GS=0.40,experimentalmaterials were divided into 7 big groups by establishing matrix[0,1].The results showed that Tibetan bred barley with the same source area were narrow genetic basis,otherwise Tibetan bred barley with the different source area were genetic difference;in our future barley breeding work,it is necessary to explore the wild relatives of barley resource and the differentgeographical origin of bred barley varieties.Strengthen the exchange among resources,and increase the genetic diversity of breedingmaterials and new varieties.
Tibetan hulless barley;Germplasm;Genetic diversity;ISJprimer
2013-07-21
973计划前期研究专项(2011CB111512;2012CB723006);国家科技支撑计划(2012BAD03B01)。
曾兴权(1975-),男,博士,副研究员。主要从事青稞遗传育种工作。
