稳定和提高牛奶蛋白质含量的营养策略
2013-02-20刘建新刘红云金晓露
■刘建新 刘红云 朱 雯 金晓露
(浙江大学奶业科学研究所,教育部动物分子营养学重点实验室,浙江杭州 310058)
牛奶及乳制品是高质量蛋白、能量和必需微量元素和维生素的重要来源,其所含的氨基酸在人类营养与动物营养中具有独特的功效。此外,牛奶还含有各种具有生物活性的蛋白质,从抗菌、促生长、提高免疫力,到具有神经内分泌活性等。乳蛋白可以分为酪蛋白(α-、β-、-κ和γ-酪蛋白等)和乳清蛋白(包括α-乳白蛋白和β-乳球蛋白)两大类,酪蛋白占乳蛋白的80%。考虑牛奶生产者和消费者的共同需求,必须同时考虑乳蛋白的含量和产量。
奶业发达国家饲料资源丰富,饲养管理水平较高。如美国原料奶的乳脂肪含量平均达3.5%,乳蛋白含量可达3.2%。我国奶牛近10多年来发展很快,但一直以数量扩张型发展,优质粗饲料供给不足,绝大多数地区以农作物秸秆作为奶牛的主要粗饲料源,奶牛平均单产不高,乳蛋白和乳脂肪含量也较低。我国最新版的生乳国家标准(卫生部,2010)中,乳蛋白、乳脂肪含量降为2.8%、3.1%,与发达国家的标准以及荷斯坦奶牛的生产潜力差距很大。
要提高牛奶的蛋白质含量,其主要策略包括育种技术和营养调控技术。由于育种手段周期漫长,所以营养调控就成为改善牛奶蛋白质含量的有效手段。在乳蛋白合成调控方面,研究主要集中在日粮氮向乳蛋白的转化效率、影响微生物蛋白合成的因素、过瘤胃蛋白对乳蛋白合成的影响,以及乳腺内蛋白质、氨基酸(包括小肽)的代谢和利用机制等方面。有研究发现,日粮蛋白质、过瘤胃蛋白质的含量和数量均可影响瘤胃微生物蛋白质和乳蛋白合成量(Choi等,2009;Broderik等,2009)。但目前仍未确定瘤胃微生物蛋白质、饲料过瘤胃蛋白质和机体内源蛋白质对乳蛋白前体物的贡献率。而且,日粮氨基酸供应及其平衡性,小肽对乳蛋白合成的贡献等也未明了。本文将对乳腺蛋白质的生物合成、改善乳蛋白合成的营养策略等方面的研究进展予以综述。
1 乳蛋白质的生物合成
乳腺的新陈代谢水平较高,乳腺合成的蛋白质占全身蛋白合成量的43%(Thivierge等,2002)。从血液中获取的游离氨基酸是牛奶蛋白质的前体物质,小肽也可能是牛奶蛋白质合成的前体物质之一(Backwell等,1996)。奶牛乳腺内蛋白质的合成与分泌是一个复杂的生物过程,包括AA摄取、DNA转录、mRNA翻译,以及乳蛋白质前体修饰直到分泌。所有的乳腺上皮细胞被认为都参与和促进这一生物过程。牛奶蛋白质含量的变化可能与乳腺内影响蛋白质代谢的营养因子(氨基酸和葡萄糖可利用性)、内分泌因子和转录下游调控因子等有关(Bobe等,2009;Burgos等,2010)。最近的研究表明,蛋白质的转录调控对于奶牛牛奶蛋白质产量可能比较重要,转录起始和延伸因子可能是乳产量调控乃至乳蛋白合成调控的潜在靶标。在采用营养策略之前,人们必须更清楚地了解控制乳蛋白质合成的机制。
1.1 乳腺
乳腺能够调节养分的吸收以便维持乳汁的合成,这表现在调节乳腺血液流动或者调节动脉供应中牛奶前体细胞的去除,此功能已得以证实(Thivierge等,2002)。乳房可以控制调节牛奶合成的胞内控件,其中包括一种影响牛奶产量的蛋白质,它是泌乳的反馈抑制剂 (Wilde等,1995)。这种反馈抑制物对牛奶蛋白分泌的调节作用是通过增加新合成的酪蛋白的降解和下调蛋白质的合成而实现(Rennison等,1993)。
1.2 内分泌
相比于其他生理系统,内分泌系统在乳腺发育、乳的生成和泌乳维持等各个方面发挥着关键作用。泌乳过程中重要的神经内分泌激素包括催乳相关激素、生长激素、胰高血糖素等。
体外培养证实,催乳相关的激素(如催乳素、皮质醇和胰岛素)对于乳腺组织和细胞分化是必需的(赵珂,2011);乳蛋白基因表达的诱导需要补充催乳素、皮质醇和胰岛素。催乳素属于多肽激素,对于分娩乃至产后乳腺发育至正常泌乳起着重要作用(Oakes等,2008),也可能在转录阶段促进乳蛋白合成(Nardacci等,1978)。皮质醇属于糖皮质激素,可增加由催乳素诱导的酪蛋白mRNA的积累(Kabotyanski等,2009),通过间接细胞机制诱导酪蛋白基因表达;添加皮质醇可以直接激发乳清蛋白mRNA的快速积累(Houdebine等,1991)。胰岛素对牛奶蛋白质合成也起着重要作用,乳腺上皮细胞中胰岛素在多个水平(牛奶蛋白质基因表达、酪蛋白合成和14C-赖氨酸的摄取)激发乳蛋白质合成(Menzies等,2009)。此外,胰岛素和催乳素可协同促进乳腺组织酪蛋白mRNA的积累(Choi等,2004)。这些激素可能通过调控哺乳动物雷帕霉素靶蛋白(mTOR)信号通路来调控乳腺蛋白质合成(Yang等,2008)。
奶牛乳腺内生长激素有促进乳汁生成的作用,但生长激素对于调控乳蛋白质合成的分子机制仍然未知。一些研究表明,生长激素的促乳生成作用与乳腺上皮细胞增殖和活性增强有关(Mo⁃lento等,2002),包括生长激素对乳腺的直接作用、通过增加IGF-1分泌而产生的间接作用(Akers等,2000)。Hayashi等(2009)研究发现,生长激素对于奶产量的作用至少部分是通过上调蛋白质翻译启动和延伸阶段来调节的;他们同时发现,生长激素处理组奶牛乳蛋白产量的增加与核糖体蛋白S6(rpS6)的磷酸化程度增加有关,这意味着生长激素对泌乳的作用可能是通过mTOR通路调控的。
在不同情况下,对泌乳早期奶牛静注胰高血糖素会导致蛋白质产量和含量短期内快速下降,而对其他乳成分或乳产量影响较小(Bobe等,2009)。最有可能的机制是,胰高血糖素通过增加肝脏内氨基酸糖原异生为葡萄糖,从而减少动脉中糖原异生氨基酸的浓度,增加葡萄糖浓度,导致乳腺氨基酸的可利用性降低,而乳糖的可利用率增加(Bobe等,2009)。
1.3 营养
蛋白质合成需要氨基酸和能源(ATP),因此提供充足的氨基酸和能源对于满足泌乳需求必不可少。不少试验用乳腺组织培养研究了氨基酸供应模式对蛋白质代谢的影响(吴慧慧,2007;来金良,2006)。添加酪蛋白可提高泌乳奶牛的乳蛋白含量和产量(Raggio等,2006);乳蛋白产量的提高与乳腺组织必需氨基酸摄取的增加有关,而且已证实某些氨基酸对乳蛋白的合成是限制性的(John⁃ston等,2004)。氨基酸除了作为乳蛋白合成的基本结构单元外,还能起到信号分子的作用。现在人们对于氨基酸作为信号分子调控蛋白质合成的作用之认识有所增加。随着氨基酸可利用性的增加,乳腺细胞中mTOR信号蛋白的活性也会提高,而且催乳激素可增强这种反应(Burgos等,2010)。
泌乳时乳腺需要充足的能量供应。Hanigan等(2009)估算出在泌乳时乳腺组织形成的ATP有近一半用来合成蛋白质。据报道,饲料模式会影响泌乳牛的奶产量和乳蛋白产量(Rius等,2010a;Raggio等,2006;Zhu等,2012)。细胞内的能量可利用性在调节蛋白质合成中起到一定作用,能源物质不仅可以用作底物,而且可以通过直接对蛋白质合成系统进行信号调节而起作用(Proud,2007)。Rius等(2010b)还证明,灌注淀粉在改变乳蛋白质产量的同时,也可相应增强rpS6磷酸化和mTOR信号通路。
2 日粮氨基酸供应与乳蛋白合成
日粮蛋白质水平是很重要的营养指标,尤其对泌乳早期奶牛,因为这个阶段奶牛的蛋白质需求量高,而干物质采食量相对较低。反刍动物营养中,日粮蛋白质在瘤胃中的降解可造成蛋白质的浪费,因此希望通过提高日粮蛋白质水平以满足奶牛对高蛋白质的需求,并不总是有效的解决策略。蛋白质营养的本质是为动物提供氨基酸,氨基酸只有在充足且均衡供应的条件下,才能被动物有效地吸收和利用。随着生物技术的发展,饲用氨基酸已实现了产业化生产。游离氨基酸是奶牛乳腺组织吸收氨态氮的主要形式,但有时乳腺组织摄取的必需氨基酸并不能满足奶牛最大合成乳蛋白的需要(Bick⁃erstaffe等,1974;Metcalf等,1996),所以奶牛乳腺组织需要从肽结合氨基酸(Peptide-bounded amino acid,PBAA)代谢池中摄取氨基酸。
2.1 氨基酸需要量的预测
氨基酸的供给和需求量是预测奶牛乳蛋白产量所必需的。Lys和Met是奶牛集约化饲养条件下的限制性氨基酸,代谢蛋白质中Lys和Met的浓度分别为7.2%和2.4%,两者之比为3∶1时,可获得最佳的乳蛋白含量。但Socha等(2005)研究表明,代谢蛋白质中Lys与Met之比为3.8∶1或3.9∶1时可获得最高的乳蛋白含量和产量。一般而言,对于奶牛来说,Met比Lys更具限制性,给奶牛饲喂过瘤胃保护Met比饲喂过瘤胃保护Lys效果要好。
消化道氨基酸被吸收进入肝脏后,会发生有差别的净代谢,其范围从支链氨基酸的0%到苯丙氨酸的50%不等(Lapierre等,2006),这个过程会改变乳腺氨基酸的供应模式。氨基酸在十二指肠到乳腺间的中间代谢会改变被吸收氨基酸合成乳蛋白的效率,因此这种转运效率应当在预测模型中予以考虑。提出准确的预测模型其目的是为了更好地估测各种因素对乳蛋白分泌的影响。Ipharraguerre等(2005)用Meta分析综合评估了不同日粮蛋白来源对到达小肠非氨态氮的影响,发现当日粮RUP含量较高时,流入十二指肠的微生物蛋白量平均被抑制7%,部分抵消了通过RUP源提高代谢蛋白总量带来的效果。尽管如此,提高RUP含量可以增加流入十二指肠的Lys或Met量,足以弥补代谢蛋白质合成减少所带来的损失。对氨基酸代谢整体知识的了解,将会帮助人们构建数学模型,以准确地定义日粮蛋白质供应对乳蛋白合成的影响。
2.2 氨基酸的供应与平衡
2.2.1 氨基酸的供应
增加氨基酸对乳腺细胞的供应是通过日粮策略提高乳蛋白含量或者产量的基础。日粮中缺乏玉米或粗料过多,或者以豆粕作为RUP源,或者两种情况并存时,Met将成为第一限制性氨基酸(NRC,2001)。补充过瘤胃保护Met从而增加Met的供应,能提高高产奶牛的乳蛋白产量,可能是由于增加了乳蛋白的合成(NRC,2001)。奶牛日粮中添加过瘤胃保护Met可以增加乳中真蛋白的含量和产量(Patton,2010)。许多研究测定分析各种Met衍生物和类似物的抗瘤胃降解特性,其中以Met羟基类似物(HMB)研究最多。添加HMB可以增加乳产量、乳蛋白产量和乳脂含量(Wang等,2010)。但St-Pierre等(2005)研究发现,HMB并不能有效满足泌乳奶牛的Met需求,至少不能满足乳蛋白合成的需求。也有报道证明,异丙酯HMB(HMBi)可为奶牛提供足够的Met(Graulet等,2005),增加奶牛的乳蛋白产量和含量(Rulquin等,2006;见表1)。据计算,HMBi中Met的生物效价是Smartamine M(SmM)的61%(Rulquin等,2006)。
Lys是奶牛的另一限制性氨基酸。增加日粮Lys和Met的营养水平,可增加奶牛干物质采食量、奶产量、乳蛋白的产量和含量(Socha等,2005)。必需氨基酸中支链氨基酸(包括亮氨酸、异亮氨酸、缬氨酸)具有某些独特的性质。支链氨基酸在调控氨基酸和蛋白代谢中起重要的作用,包括促进胰岛素分泌、正向调控蛋白合成、作为谷氨酰胺合成的供体和器官之间的信号传递者(Lal等,1995)。支链氨基酸对奶牛泌乳也起重要作用,可占乳蛋白中总氨基酸的20%,或占必需氨基酸的50%(Jenness,1974),而且支链氨基酸参与糖酵解和三羧酸循环,形成的中间体可进入非必需氨基酸代谢库,用于乳蛋白的合成所需(Mepham,1982)。

表1 补充不同形式的蛋氨酸对奶牛泌乳性能和乳蛋白产量的影响
2.2.2氨基酸平衡
日粮某些AA过剩或不足可导致AA不平衡,会限制奶牛乳腺蛋白质的合成,并降低其泌乳性能。研究发现,酪蛋白αs1基因在体外培养的奶牛乳腺上皮细胞中的表达需要每种必需氨基酸(Met、Lys、Phe和Thr)都处于最适浓度,任一必需氨基酸浓度过高或过低都会降低乳蛋白基因的表达(Zhao等,2005;Zhou等,2008)。短期饲养试验结果表明,当奶牛以青贮牧草和谷物为基础日粮,以羽毛粉作唯一的蛋白源时,奶产量因特定AA,如His、Met和Lys的缺乏而下降(Kim等,1999、2000)。给泌乳早中期和中晚期奶牛饲喂缺乏特定AA的日粮6周,发现奶产量明显下降(Yeo等,2003)。Wang等(2010)评价了可代谢蛋白质中Lys和Met的比例对中国荷斯坦奶牛泌乳性能和AA代谢的影响,发现补充Met和Lys,Met或Lys可增加乳腺对AA的吸收;补充Lys或Met增加了奶产量,但对乳成分没有显著影响;Lys和Met为最佳比例3∶1时,乳蛋白含量和奶产量达最大值。
改善可吸收AA的平衡性可提高氨基酸在乳蛋白合成中的利用效率。已经证实,提高Lys和Met营养能降低用于获得相同乳蛋白产量的日粮粗蛋白需要量(Rulquin等,1990)。早期泌乳奶牛易受肠道中AA平衡的影响,优化日粮中Lys和Met的营养可以提高其泌乳性能(Socha等,2005;见表2)。这些现象可用AA限制理论得到解释,即在给定的饮食或者生理条件下,仅有一种AA吸收供给受限也会影响奶牛乳蛋白产量(Weekes等,2006)。用乳腺组织体外培养发现,Lys与Met比例、Thr与Phe比例都会影响乳腺中αs1酪蛋白基因的表达,它们的最佳比例分别为3.5∶1(Liu等,2007)和1.05∶1(Zhou等,2008)。在奶牛试验中我们还发现,在补充Lys、Met、Thr和Phe后,即使降低日粮蛋白质含量,其奶产量不减反增,其饲料N用于泌乳的效率从对照组的25%提高到36%(杨志强,2009;见表3)。由此可得出结论,适宜的必需AA比例可促进乳蛋白的合成,改善AA平衡有利于提高乳蛋白产量和奶牛生产性能。

表2 两个蛋白水平下补充瘤胃保护蛋氨酸对奶牛105 d泌乳性能的影响

表3 饲喂较低蛋白含量但补充氨基酸日粮时奶牛的生产性能与氮利用率
2.2.3 AA在调控蛋白质合成转化机制中的作用
调控奶牛产奶量和乳蛋白质产量的内在分子机制尚未完全清楚。Hayashi等(2009)指出,mTOR通路可能对乳腺中乳蛋白合成具有潜在调控作用。AA不仅是营养物质,还可能是信号分子。人们日益认识到AA在蛋白质合成中的信号分子作用(Kimball等,2002)。日粮中AA可以通过mTOR信号转导通路调控动物体内mRNA翻译起始阶段,mTOR是控制核糖体合成的蛋白激酶 (Jeffer⁃son等,2003)。Moshel等(2006)研究了AA对催乳素条件下体外培养的乳腺上皮细胞翻译水平调控中的作用,发现去除总AA或选择性去除亮氨酸对牛乳腺细胞β-乳球蛋白合成有负面影响,AA参与翻译的直接信号作用决定乳腺上皮细胞牛奶蛋白质的合成效率。
2.3 肽的供应
游离氨基酸是泌乳期乳腺组织吸收氨态氮的主要形式,这一概念长期以来一直遭到质疑。对于泌乳奶牛来说,通过乳腺吸收的特定必需氨基酸往往不能满足乳腺分泌蛋白质的需要(Bicker⁃staffe等,1974;Metcalf等,1996)。当氨基酸不能满足乳蛋白合成的需要时似乎加快自身从肽结合氨基酸池中的回收。
Tagari等(2004)研究发现,给泌乳奶牛无论是饲喂蒸汽压片玉米或压碎玉米的日粮,均发现大量的肽结合氨基酸从门静脉流出。使用15N或14C特殊标记技术在泌乳奶山羊体内的研究发现,许多必需氨基酸是以肽结合氨基酸的形式被乳腺组织回收用于合成蛋白质(Bequette等,1999)。尽管在血细胞中储备大量的Lys,从游离氨基酸池中摄取的Lys并不能满足乳蛋白质合成的需要(Hanigan等,1991),其中很大一部分是从肽结合Lys池中摄取的,再次支持了肽结合氨基酸池作为第一储备氨基酸源的观点。我们的研究也发现,当用蛋氨酸二肽替代Met时,αs1酪蛋白基因在乳腺上皮细胞中表达量增加(Wu等,2007;见图1和图2)。同样,相比游离氨基酸,用苯丙氨酸二肽替代苯丙氨酸促进了αs1酪蛋白基因乳腺上皮细胞中的表达(Zhou等,2011,见图3)。这些结果与组氨酸二肽相比游离组氨酸增加乳蛋白产量(Backwell等,1996)的报道一致。Tagari等(2008)报道,肽结合氨基酸是乳腺摄取总游离氨基酸的一部分,占3.7%~4.8%,但是个别肽结合必需氨基酸占2.5%~23.8%。
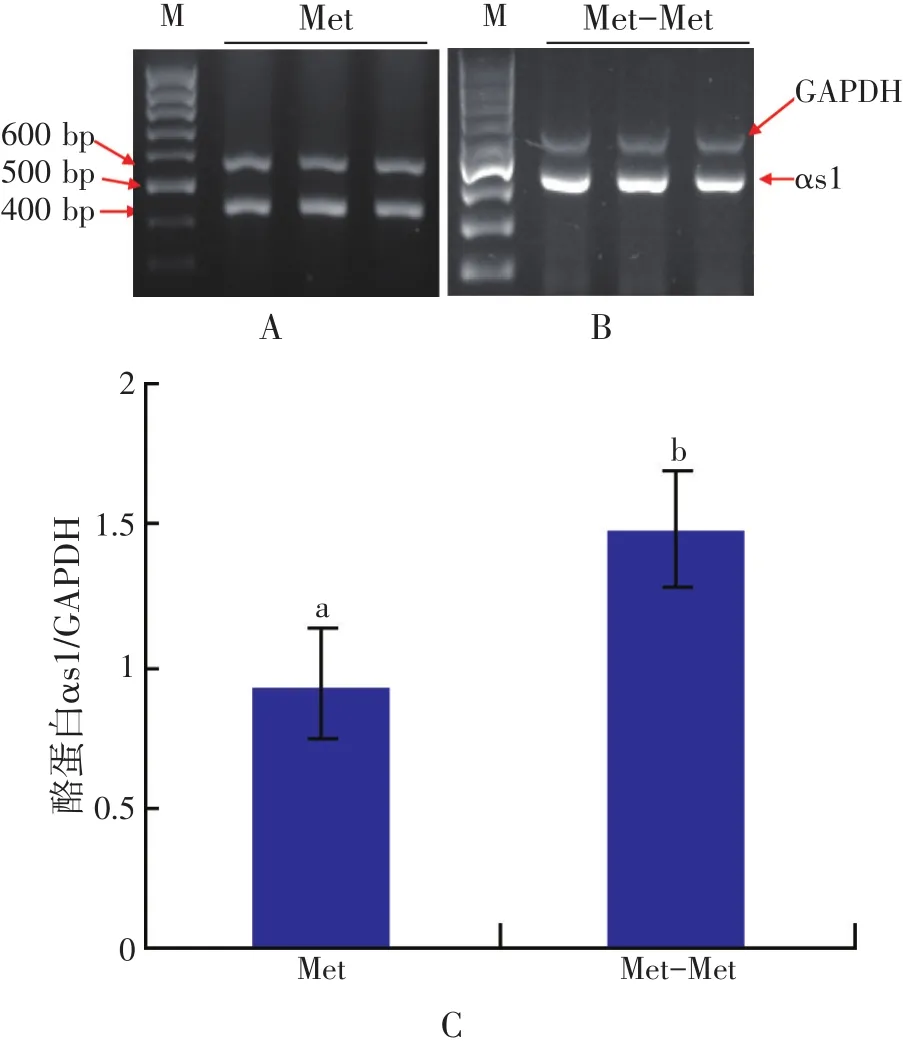
图1 蛋氨酸二肽和蛋氨酸对酪蛋白αs1基因表达的影响
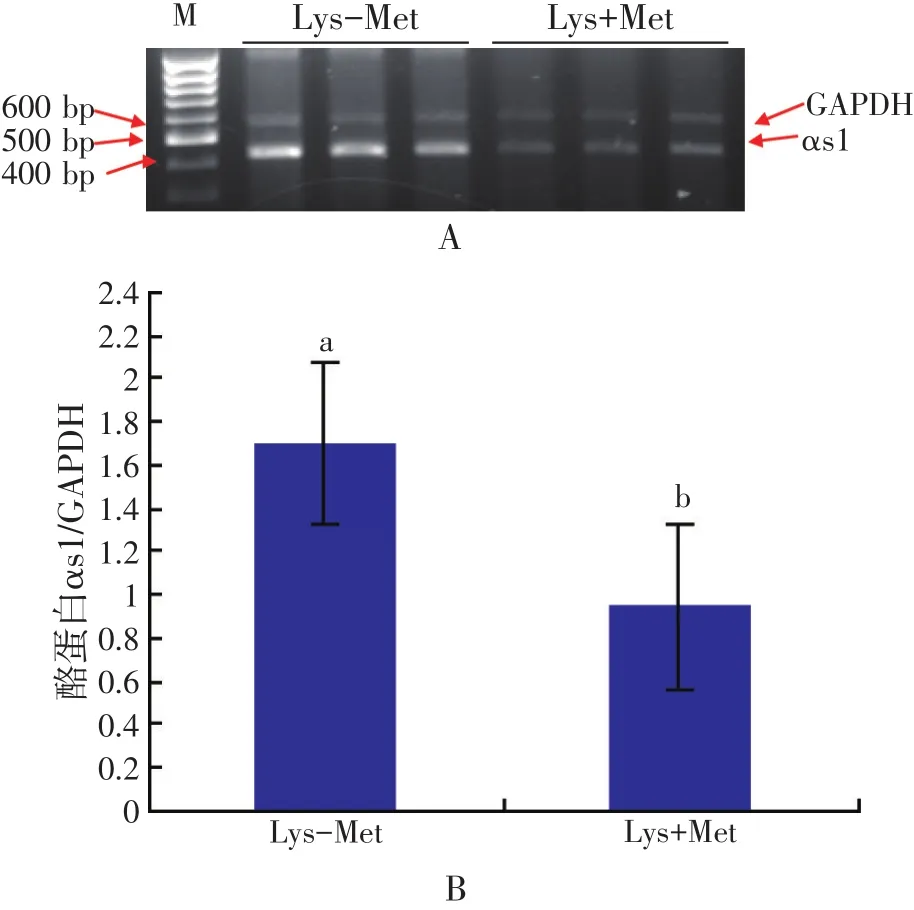
图2 蛋氨酸赖氨酸二肽和蛋氨酸、赖氨酸对酪蛋白αs1基因表达影响比较
乳腺上皮细胞吸收完整的寡肽的分子基础是位于顶端的H+/肽共同转运载体,肽转运载体1(PepT1)和肽转运载体2(PepT2)。高容量低亲和力的肽转运载体1(PepT1)主要在小肠部位表达(Adibi,1997),奶牛乳腺中没有发现PepT1 mRNA的表达(Zhou等,2011)。高亲和力低容量的肽转运载体2(PepT2)主要在肾小管处表达(Shen等,1999)。用反转录PCR方法检测到PepT2 mRNA在小鼠乳腺细胞提取物、人乳腺上皮细胞(Groneberg等,2002)和奶牛乳腺组织块中均有表达(Zhou等,2011)。乳腺肽转运载体的发现可能会为蛋白质代谢和分泌的研究提供新的视角。

图3 苯丙氨酸寡肽对体外培养乳腺上皮细胞αs1酪蛋白基因表达的影响
3 日粮模式与乳蛋白合成
增加代谢蛋白质的供应可增加奶产量和乳蛋白产量,在4%FCM和乳蛋白产量与日粮代谢蛋白质含量之间存在非线性回归(Wang等,2007)。代谢蛋白质主要来源于饲料过瘤胃蛋白和瘤胃合成的微生物蛋白两部分,其中微生物蛋白占可吸收蛋白质的50%~80%(Storm等,1983)。由于饲料蛋白资源短缺,最大限度地发挥瘤胃微生物功能,合成微生物蛋白具有十分重要的意义。除了瘤胃可降解氮之外,可利用能对于瘤胃微生物蛋白的合成至关重要。
3.1 饲料组合
常规奶牛日粮配制中,粗饲料占40%~50%,不仅影响DMI,同时影响能量采食量(Kendall等,2009)。发达国家利用苜蓿、全株玉米等优质粗饲料,奶牛生产水平、牛奶质量都较高。我国这些资源都比较匮乏,近年来大量进口苜蓿等优质牧草,如2011年进口27.6万吨,这种局面难以持久。我国现行养殖模式是利用大量农作物秸秆(玉米秸等),同时配以大量精料,导致奶牛瘤胃代谢异常,产生健康隐患,影响牛奶产量和质量。在我国现行养殖模式下,如何改善牛奶质量,是重要的研究课题。
日粮粗饲料组合对瘤胃微生物蛋白合成的影响,不同研究的结果并不一致(Voelker Linton等,2009;Khorasani等,2001)。Clark 等(1992)总结 28个试验的结果发现,瘤胃内有机物降解是影响微生物蛋白合成的重要因素。Meta分析结果也表明,非结构碳水化合物含量越高,奶牛的微生物蛋白产量也越高(Nocek等,1988)。也有报道认为,可利用碳水化合物的低利用率与微生物蛋白质产量有负面影响。Zhu等(2012)的试验研究表明,奶产量与微生物蛋白产量有显著的回归关系,表明在玉米秸等作物秸秆用于奶牛日粮时,对促进瘤胃微生物蛋白产量至关重要。
在最近完成的一项研究(Zhu等,2012;见表4)中,我们以玉米秸、羊草、苜蓿干草配制不同NFC/NDF比例的日粮饲喂奶牛,发现苜蓿日粮比玉米秸日粮含有较高的可溶性糖和淀粉,能提供较多的瘤胃易发酵碳水化合物,从而生成较多的瘤胃MCP(1 876 g/d∶1 601 g/d),贡献较多的乳蛋白前体物——小肠代谢蛋白,提高乳蛋白含量和产量。换言之,饲喂玉米秸日粮产生较低的微生物蛋白,可能与日粮中可溶性糖分不足,降低了MCP合成效率密切相关。
3.2 补充糖分和添加AA
在前述研究的基础上,我们进一步开展了补充能源及氨基酸的研究(朱雯等,未发表)。以泌乳奶牛为对象,以苜蓿和青贮玉米型日粮为对照,在玉米秸日粮中补充淀粉,并添加必需AA(Lys、Met),研究发现(见表5),仅补充淀粉时,在产奶量、乳蛋白含量和产量、氮利用效率上均略低于苜蓿日粮,但差异不显著;而同时补充适量淀粉和必需AA后,奶牛的DMI、产奶量、乳成分和氮利用效率与苜蓿日粮无明显差异,表明以玉米秸为主要结构性碳水化合物源时,需同时补充易发酵糖分和必需AA,才能达到最佳效果。该研究结果再次证明,氨基酸对于乳蛋白合成的重要性和必要性。
4 结语
奶牛能够为人类提供丰富的乳蛋白,改善人类健康。蛋白质在奶牛乳腺内的合成是十分复杂的生物学过程,受到营养、内分泌和其他众多调控因子的影响。增加乳腺的氨基酸供应是增加牛奶蛋白质含量和产量的关键措施,足够而平衡的氨基酸供应对于奶牛的泌乳性能和乳蛋白合成至关重要。整合氨基酸代谢知识,改善氨基酸需要量的预测等将有助于建立蛋白质/氨基酸需要量模型,以准确地提供日粮蛋白质。需要进一步研究泌乳奶牛的氨基酸代谢,包括氨基酸、小肽用于合成乳蛋白的效率等。同时需要研究以确定奶牛的限制性氨基酸、不同氨基酸或小肽对乳蛋白合成的影响,乳腺对肽结合氨基酸的利用,以及环境、营养和不同生理状态下乳蛋白的合成。
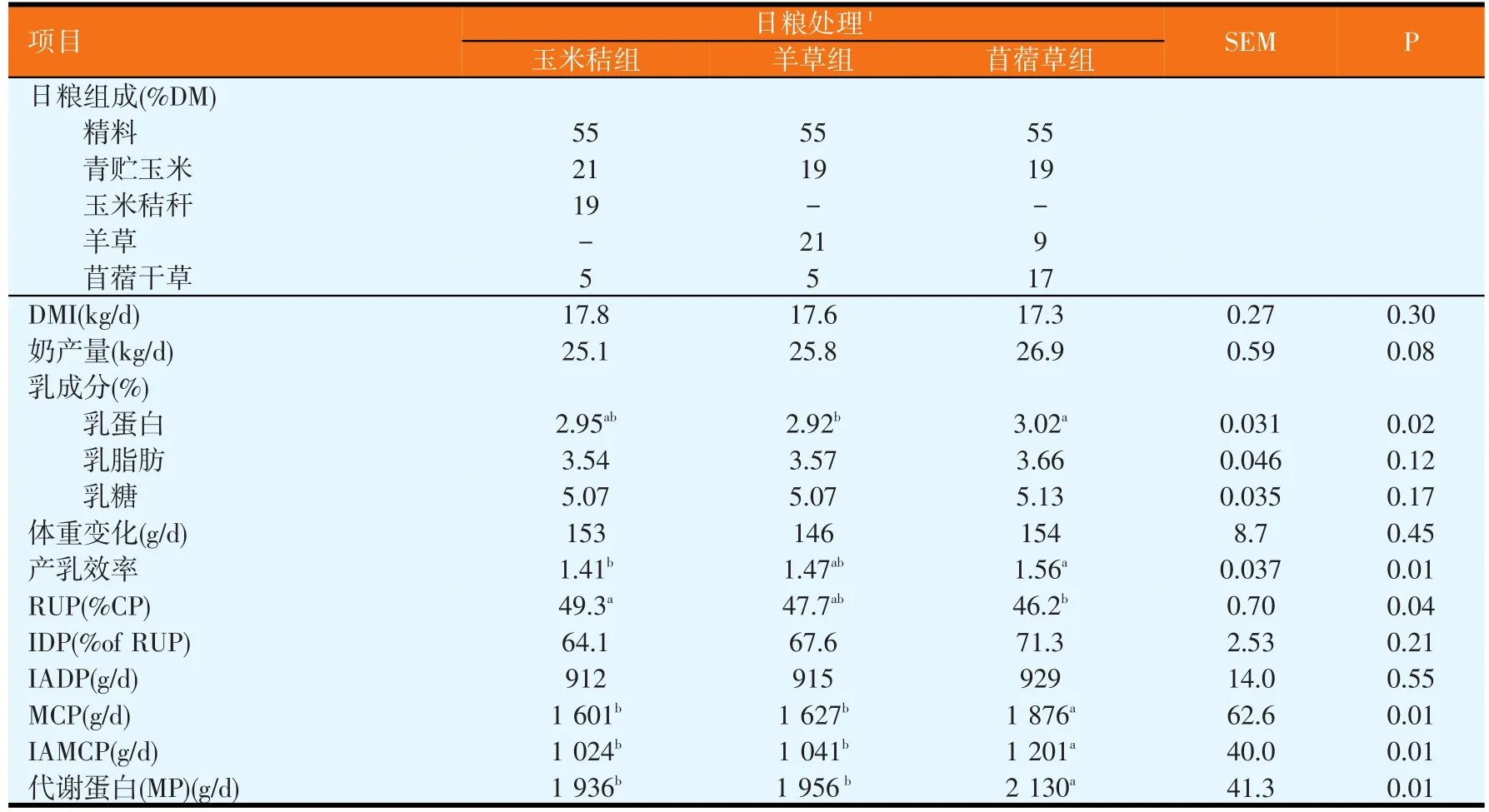
表4 玉米秸、羊草和苜蓿干草日粮的奶牛干物质采食量和奶产量之比较

表5 玉米秸、羊草和苜蓿干草日粮的奶牛干物质采食量和奶产量之比较
