山茱萸种子休眠机理与解除方法初探
2013-01-05洑香香周晓东刘红娜
洑香香,周晓东,刘红娜
(南京林业大学 森环院,江苏 南京 210037)
山茱萸种子休眠机理与解除方法初探
洑香香,周晓东,刘红娜
(南京林业大学 森环院,江苏 南京 210037)
山茱萸种子具深休眠特性,在自然条件下需2年才开始萌发。本研究用500 mg·L-1GA3分别浸泡山茱萸种子3 d和5 d,结合不同层积处理(变温(5~30℃)层积)、低温层积(0~5℃)、低温层积1个半月后转入变温层积),以探索解除种子休眠的方法;并对层积前后的种子进行解剖比较研究。结果表明:GA3和层积处理都可促进种子休眠的解除;层积3个月的结果表明变温层积解除休眠的效果最好,低温层积+变温层积次之,低温层积无效果,但低温层积7个月可使发芽率达到40%;GA3浸泡结合层积处理可有效解除种子休眠。处理前后种子的解剖结构变化表明:种子的休眠原因既有种皮的机械束缚作用(物理休眠),同时还存在胚的形态休眠。因此山茱萸种子的休眠属于复杂的多因素综合休眠。
山茱萸;种子休眠;GA3;层积处理; 解剖结构
山茱萸Cornus officinalis具有极好的观赏性,作为一种新的彩色美化树种被逐步认可。山茱萸先花后叶,花色金黄,似腊梅、迎春和连翘;花期1~3月份:长江以南地区在春节前后,黄河、淮河及以北地区在春节期间,北京及以北地区在元宵节期间即次第开放;花期长达50 d,料峭春寒,分外妖娆。因山茱萸具有花期早、花期长、花干性耐寒的特点,是极具市场开发潜力的年宵花;秋果红色至紫红色,簇果如珠,绯红欲滴,艳丽悦目,是秋冬著名的观果佳品。
山茱萸属生长缓慢、结果迟、寿命长的树种。实生苗在天然状态下7~10年才能开花结实,人工栽培条件下5~6年可正常开花但不结实,8~9年后多数植株挂果[1]。种子具深休眠习性,需层积一年以上才能大量萌发[2]。由于山茱萸繁殖能力的限制,严重影响了其在园林市场的推广和应用。本研究对种子采用不同的处理方法,旨在短时间打破休眠并初步探讨种子休眠的原因,为生产上规模化育苗提供技术支撑。
1 材料与方法
1.1 试验材料
种子材料来自于安徽黄山,果实采收于2010年10~11月份,采收后搓去果肉,淘洗干净后晾干,于0~4℃下湿沙保存。
1.2 试验方法
1.2.1 种子处理
层积前将种子用浓度为500 mg·L-1GA3在25℃条件下分别浸泡3 d和5 d,每天换液一次;对照用蒸馏水浸泡。将预处理的种子与湿沙以1∶3进行混合并进行层积处理。层积处理设3个温度:变温层积(5~30℃)、低温层积(0~5℃)、低温层积1个半月后转入变温层积(见表1)。层积期间管理注意保持通气,并用500 mg·L-1GA3溶液浇灌、对照用水浇灌,以保持湿润。

表1 山茱萸种子处理试验Table 1 Processing test of C. officinalis seed dormancy
1.2.2 发芽测定
层积处理至种子开始露白时,取样测定发芽率。发芽测定采用3个重复,每个重复50粒种子。发芽条件:种子置于沙床在光照培养箱中进行,发芽温度为20~30℃,光照为白天8 h。种子发芽标准参照《林木种子检验规程》(GB 2772-1999)的规定,每隔3 d计数一次,发芽持续时间为1个月。
发芽率:在规定发芽天数内正常发芽的种子粒数占供试种子粒数的百分比;
发芽势:日发芽粒数达到高峰时正常发芽种子数占供检种子数的百分率。
发芽持续时间:根据《林木种子检验规程》(GB 2772-1999)[3]的规定,以连续3天每天的发芽率不超过重复粒数的1%为结束发芽。
1.3 数据统计分析
统计种子发芽率、发芽势,采用Excel软件对测定数据进行方差分析。
2 结果与分析
2.1 山茱萸种子萌发特性
2.1.1 种子的萌发进程
以用500 mg·L-1GA3浸泡5 d、并置于低温条件下层积7个月来阐述种子的发芽进程。
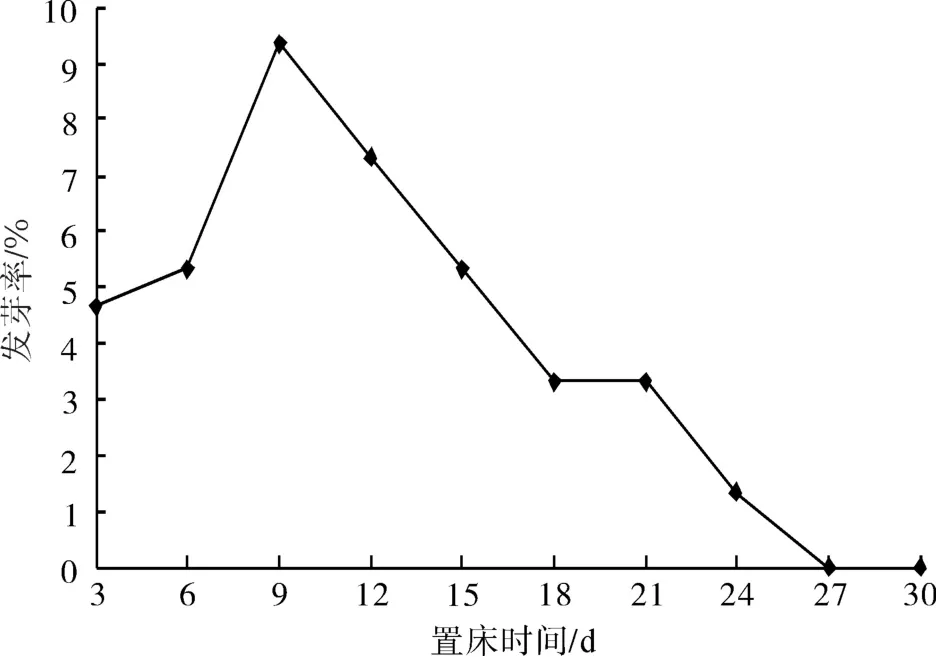
图1 山茱萸种子萌发进程Fig.1 Germination process of C. officinalis seeds
由图1可知:在持续1个月的发芽时间内,合计发芽率为40%。置床后第9 d出现了发芽高峰(发芽率达到9%),发芽势为19%。萌发过程中发芽情况相对平稳,这和田间表现一致,即在田间发芽持续时间可长达6个月。由此可知,山茱萸种子由于其深休眠特性,解除休眠也较为缓慢,在萌发过程中没有明显的发芽高峰。
一般认为,发芽势越高种子活力强,发芽迅速、整齐,幼苗生产潜力大,幼苗间个体差异小,并有利于田间管理。但是山茱萸种子的发芽势在本试验中最高仅达19%,可能由于种子未完全解除休眠,因而种子发芽率和发芽势均不太高。
2.2 GA3处理对种子萌发的影响
植物激素处理对解除种子休眠(尤其是生理性休眠)的效果显著。由表2可以看出(层积3个月),GA3处理可以明显促进山茱萸种子的萌发。单因素方差分析和多重比较分析表明(表2),GA3处理3 d和5 d的种子发芽率均与对照差异显著,但前两者之间差异不显著,说明GA3处理明显促进了山茱萸种子的萌发,但处理时间长短对解除休眠的效果不显著。
2.3 层积温度对解除山茱萸种子休眠的效果
层积是常用的有效打破种子休眠的方法,尤其对因含抑制物质如ABA等而形成生理休眠(PD)的种子效果显著,同时也可有效解除物理休眠(PY)类种子[4]。
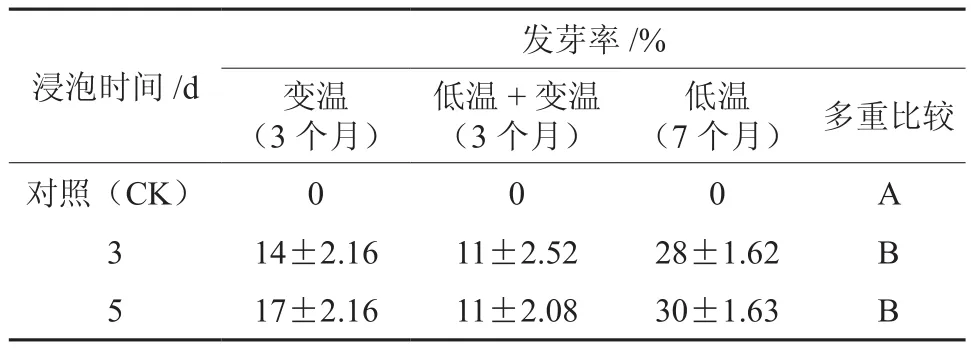
表2 GA3浸泡山茱萸种子的发芽率及比较†Table 2 Germination percentage of C. officinalis seeds treated with GA3 and stratification
由表3可知,层积处理3个月时种子发芽率差异很大。变温层积效果最好,发芽率可达36%;低温层积1个半月后转入变温层积的发芽率次之,仅达9%;而低温层积3个月种子的发芽率为0,说明种子仍处于休眠状态。
方差分析表明,不同层积处理间种子的发芽率达差异极显著水平(p<0.01)。由此可见层积处理对山茱萸种子萌发有显著的促进作用。多重比较显示低温处理、变温和低温+变温处理差异达极显著水平,而后两者间的差异达显著水平。说明变温层积在解除山茱萸种子的休眠中起了更为重要的作用,相比较而言,低温层积的作用则小一些。
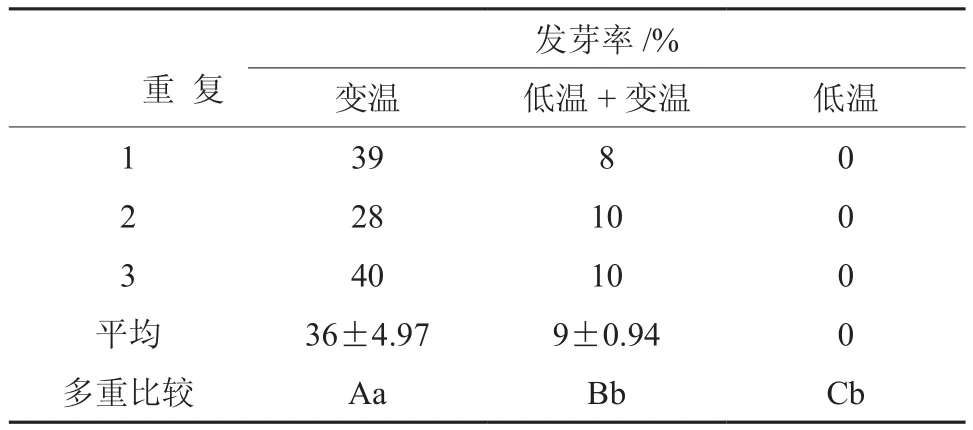
表3 层积处理3个月对山茱萸种子休眠解除的影响†Table 3 Effects of stratification for 3 months different temperatureon C. officinalis seeds releasing dormancy
2.4 GA3浸泡结合层积处理对解除种子休眠的综合影响
为了分析激素处理(时间)和层积温度对解除种子休眠的综合影响,本研究对用500 mg·L-1GA3分别浸泡3、5 d的种子在变温、低温+变温和低温条件下层积3个月进行综合分析(见表4)。由表可见,仅从发芽情况看,GA3浸泡3 d并在变温条件下层积的发芽效果最好,可达36%,但该处理在低温+变温层积的效果较差,发芽率只有12%;GA3浸泡5 d的种子在变温、低温+变温条件下层积的效果相似,发芽率都达到17%;对照(清水处理)、GA3浸泡后在低温下层积在3个月对解除休眠的效果不显著。
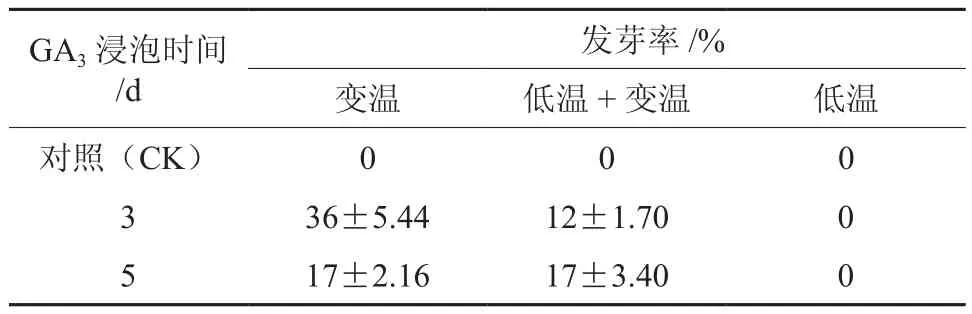
表4 GA3结合层积处理山茱萸种子的发芽率Table 4 Germination percentage of C. officinalis seeds after stratification and GA3 treatments
方差分析表明(表5),GA3处理的时间、层积方法及二者的交互作用都对种子的发芽率的影响达到了极显著水平。由此可见,GA3处理和层积方法分别在一定程度上能解除种子休眠,从交互效应的影响效果看,二者结合处理可加速解除种子的休眠。
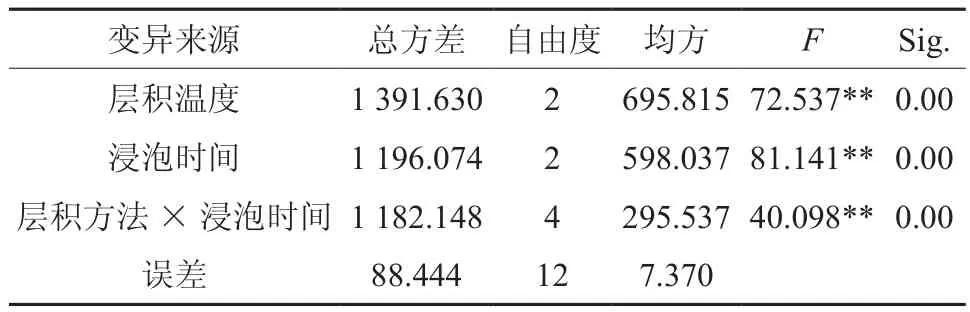
表5 预处理对种子发芽率影响方差分析Table 5 Variance analysis of effect of pretreatment on seed germination
2.5 山茱萸种子休眠原因初探
为了进一步探讨山茱萸种子的休眠原因,本研究把预处理前后的种子进行解剖比较,以期找出山茱萸种子的休眠原因。
从种子的横切图(如图2)可看出,明显可见胚、胚乳和骨质种皮结构。最为明显的是种皮厚度与整个胚腔直径相近,且种皮中镶嵌一圈蜂窝状的分泌组织,内含黄色的粘性油状物质,管康林等[2]认为这些粘性物质主要为单宁类物质。种皮主要是由角质层细胞、石细胞和上述分泌组织组成。刘雅帅[5]则发现种皮的角质层细胞和石细胞排列紧密,影响了种子的透气和透水作用;同时认为单宁类粘性油状物质对种子的萌发具有抑制作用。种子经层积处理后种皮软化,油状物质逐渐消失(解剖过程中的粘性感消失)。由此推测山茱萸种子种皮的机械障碍及种皮内抑制物质的存在是种子休眠的主要原因之一。
从横切图上(种子的1/2处切)可见,层积前种胚直径D(图2-A)约为胚腔直径的1/2,胚的横断面(推测应是胚轴的横断面)仅约占胚腔的1/4;经过层积处理后(图2-B)胚直径D约达到胚腔直径的3/4以上。从纵切图上看,层积前(图3-C)明显可见胚体的长度约为胚腔长度的1/2~3/4,而经过层积处理后胚体的长度基本上充满了整个胚腔(图3-D),由此可见,山茱萸种子还存在胚的形态休眠,即种子成熟时,胚还未完成生长;经过GA3浸泡的种子在层积过程中种胚长大,从而完全生理后熟,并促进种子萌发。

图2 层积前后种子横切面:示胚横向的变化(A:层积前,B: 层积后)Fig.2 Transverse sections of C. officinalis seeds before and after stratification ( A: before stratification; B: after stratification)
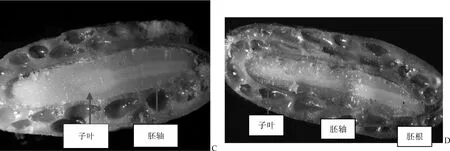
图3 层积前后种子纵切面:示胚纵向的变化(C:层积前;D:层积后)Fig.3 Longitudinal sections of C. officinalis seeds before and after stratification (C: before stratification; D: after stratification)
3 讨 论
种子休眠特性是植物对环境条件的生物学适应,对植物本身是一种保护,但也给林木种苗生产带来了困难。种子休眠过深,会导致田间出苗率降低,出苗参差不齐,同时也会造成种子的大量浪费,因此采取相应的措施解除种子休眠,具有重要的生产实践意义。
3.1 激素与休眠
对于大部分的休眠类种子中,涉及到2种激素,一是脱落酸(ABA),二是赤霉素(GA)。一般认为,在种子的成熟过程中种子内的ABA含量逐渐增加,休眠现象明显增加[6]。在成熟过程中ABA的作用是促进种子贮藏蛋白和脂肪的合成,促进种子脱水耐性和种子休眠的获得,抑制种胚萌发。GA的主要作用是解除种子休眠,促进萌发;GA在促进种子萌发时至少起到两种信号作用:第一是软化种胚周围的组织,克服种皮机械限制,第二是促进种胚的生长[7]。种子休眠或萌发取决于ABA和GA浓度的平衡,GA含量的高低决定着种子能否萌发。
GA是解除由ABA诱导的休眠的必要条件。因此对于解除由ABA引起的生理性休眠种子时,用GA处理可有效地促进种子的萌发。研究表明,用 400 mg·L-1GA3浸泡冬青(Ilex purpurea)种子48 h可显著提高GA3、细胞分裂素(CTK)和生长素含量,显著降低了ABA的含量,从而促进了萌发[8];李淑娴等[9]用500 mg·L-1GA3浸泡乌桕(Sapium sebiferum)种子24 h,可有效打破其种皮内含抑制性物质的休眠。本研究也发现,用500 mg·L-1GA3浸泡种子3~5 d,与对照相比,可大大提高种子的萌发率;对照在试验期内种子未见有萌发。
GA 通过与ABA 和环境因子的复杂作用正向调节种子休眠的解除和促进种子萌发, 由此看来在种子休眠和萌发方面ABA和GA的作用相互拮抗。但是更多的研究证实ABA /GA,而不是激素的绝对含量控制种子休眠与萌发[10-11]。对于深休眠种子,不同种源种子内的ABA浓度也相差较大,GA处理的效果则不相同。本研究对不同种源的种子进行相同处理后的发芽率表明差异显著,其中河南种源的发芽率仅有12%(资料未列出),最高的是黄山种源达40%。由此可见不同种源的种子休眠深浅存在明显差异,因此对GA的响应也存在差异。
3.2 层积与休眠
层积处理能解除种子休眠。在水合条件下,大多数非热带物种的种子经历相对低的温度(通常1~10℃的范围,一些物种为15℃)能解除休眠。自然条件下, 为了在合适的季节(通常在春季)萌发和随后的生长,水合种子的休眠在冬季通过自然层积被逐渐解除。具物理休眠和生理休眠的种子, 层积处理可有效地解除休眠。对于不同的休眠原因,层积处理解除休眠所需要的时间是不同的:解除苹果Malus pumila种子的胚休眠需40~50 d的低温层积,解除胚乳休眠需要70~80 d的低温层积处理,整个种子则需要80 d以上的低温处理[12]。通常木本植物的种子需要较长的层积处理时间, 一般为60~90 d,甚至更长;变温层积可有效解除形态休眠类种子,凌世瑜[13]研究发现解除水曲柳Fraxinus mandshurica种子休眠的最佳途径是先暖温后低温的变温层积处理。本研究也发现,变温层积比低温+变温层积更有效解除种子的休眠作用,说明变温层积首先有效促进了胚的后熟作用,同时软化种皮并降低了种皮中抑制物质的含量,从而促进种子萌发。
3.3 山茱萸种子的休眠与解除
尽管山茱萸种子存在长时间的休眠,但有关山茱萸种子休眠的研究报道却相对较少,且其休眠原因仍是一些推测性的结论,并未作系统的研究。一些研究认为山茱萸种子休眠的原因是种皮的抑制性(机械限制和抑制物质的存在)、胚未完成生理后熟或未发育完全[14]。管康林等[15]认为高温对解除种子有一定的效果,研究发现高温层积(20~28℃)3个月,再变温层积(5~17℃)3个月,可使种子发芽率达到30~50%;陈惠等[14]认为经浓硫酸处理16~17 h的种子再用500 mg·L-1GA3浸泡种子24 h后,胚根的露出率可达40%。
综合前人研究和本研究结果,认为山茱萸种子的休眠既有种皮原因(物理休眠),也存在种胚后熟问题(形态休眠)。刘雅帅[5]通过对山茱萸种子的吸水试验和种皮的扫描电镜观察得出其种皮的透水、透气性差及种皮的机械束缚是种子休眠的原因之一,有力地支持了物理休眠的存在;同时骨质种皮内的抑制性物质(粘性物质)是生理休眠的来源之一;本实验对预处理前后种子的解剖观察则提供了胚形态休眠的证据,但是否存在胚的形态生理休眠还需更深入的研究。
基于种子休眠的原因,相应的打破种子休眠的方式主要是用GA结合层积处理。从生产上来说,本研究认为目前较为有效的方法是500 mg·L-1GA3浸泡种子24 h以上并结合变温层积可有效打破种子的休眠,这和具形态休眠的银杏种子的解除方法相似[16],但该方法并未彻底解除山茱萸种子的休眠;低温层积较长时间的效果可能会更好,但当解除休眠时已错过了适宜的生产季节。因此如何在短时间内彻底打破种子休眠仍需进一步探讨。
[1] 韩东锋, 钱拴提, 朱 祥. 山茱萸幼树开花结实期调查研究[J]. 中国林副特产, 2009, 3:13-15.
[2] 管康林, 葛慧华. 山茱萸研究现状与发展[J]. 经济林研究,1990, 8(1): 14-18.
[3] 国家质量技术监督局,GB 2772-1999 林木种子检验规程[S].北京:中国标准出版社,1999.
[4] Baskin J M, Baskin C C. A classification system for seed dormancy[J]. Seed Science Research,2004,14: 1-16.
[5] 刘雅帅. 山茱萸种子休眠机理的研究[D]. 南京:南京林业大学,2008.
[6] Kushiro T, Okamoto M, Nakabayashi K, et al. The Arabidopsis cytochrome P450 CYP707A encodes ABA 80-hydroxylases: key enzymes in ABA catabolism[J]. EMBO J, 2004, 23: 1647-1656.
[7] Yamaguch I S, Kamiya Y. Gibberellins and light stimulated seed germination[J]. J Plant Growth Regul, 2002, 20: 369-376.
[8] 王 宁, 梅海军, 袁美丽, 等. 赤霉素浸种和变温层积过程对冬青种子激素含量的影响[J]. 河南农业大学学报, 2010,44(5): 524-527.
[9] 李淑娴, 刘菁菁, 田树霞, 等. 乌桕种子休眠原因及解除方法研究[J]. 南京林业大学学报, 2011, 35(5): 1-4.
[10] 张 鹏, 沈海龙. 白蜡树属树种种子休眠及其萌发的调控[J].植物生理学通讯, 2006, 42(2): 354-358.
[11] 程广有, 唐晓杰, 高红兵, 等. 东北红豆杉种子休眠机理与解除技术探讨[J]. 北京林业大学学报, 2004, 26(1): 5-9.
[12] Visser T. The role of seed coats and temperature in after-ripening,germination and respiration of apple seeds[J]. Proc K Ned Akad Wet C, 1956, 59: 211-222.
[13] 凌世瑜. 赤霉素对水曲柳种子解除休眠的作用[J]. 林业科学,1986, 22(1): 78-85.
[14] 陈 惠. 山茱萸种子休眠的破除与试管萌发[J]. 山西农业大学学报, 1996, 16(3): 262-264.
[15] 管康林, 方 星, 郑 钢. 山茱萸的休眠原因与萌发条件[J].植物生理学通讯, 1989, 5:24-27.
[16] 曹帮华,蔡春菊. 银杏种子后熟与内源激素变化的研究[J].林业科学,2006,42(2):32-37.
A preliminary study on seed dormancy mechanism and relieving methods in Cornus officinalis
FU Xiang-xiang, ZHOU Xiao-dong, LIU Hong-na
(College of Forest Resources and Environment, Nanjing Forestry University, Nanjing 210037, Jiangsu, China)
Seed of Cornus officinalis possesses deep dormancy, and begin to geminate after two years stratification. In order to reveal the mechanism of seed dormancy and to overcome seed dormancy of C. officinalis, GA3and stratification treatments were conducted.500 mg·L-1GA3solution was used to soak the seeds for 3 days and 5 days, respectively, before stratification treatments including alternative temperature at 5℃ ~30℃ , cold stratification at 0℃ ~5℃ , as well as cold stratification for 1.5 months followed by alternative temperature. At the same time, the anatomical structures of untreated and treated seeds were observed to compare the changing profiles.The results showed, in some degree, both GA3soaking and stratification treatment could overcome seed dormancy. Seed germination percentages for stratified-treatment for 3 months showed the stratification treatment at alternative temperature played more effective role than cold stratification for 1.5 months followed by alternative temperature on releasing seed dormancy, and it seemed that treatment of cold stratification for short period (≤3 months) had no effect. However, the germination percentage was up to 40% when seeds were treated with cold stratification for 7 months. So, GA3soaking combined stratification is the effective way to release the seed dormancy of C. officinalis. Seed dormancy is supposed to derive from both physical dormancy of seed coat and morphological dormancy of embryo based on comparative observation on anatomical structure of treated and control seeds. In short, the seed dormancy of C. officinalis could be classified into comprehensive dormancy caused by coat and undeveloped embryo.
Cornus officinalis; seed dormancy; GA3; stratification; anatomical structure
S722.1
A
1673-923X(2013)04-0007-06
2012-08-07
国家林业局948项目“北美四照花类观赏种质资源与培育技术引进”(项目编号:2012-4-60)
洑香香(1969-),女,教授,博士,研究方向:人工林定向培育、种子生物学;Email: xxfu@njfu.edu.cn
[本文编校:欧阳钦]
