GDF5 基因rs224331 多态性与汉族青少年体格发育指标关联分析
2012-12-23李晓静王健民高玲玲沈水仙支涤静忠程若倩奚立叶赵诸慧郑章乾章淼滢罗飞宏
裴 舟 李晓静 王健民 杨 毅 高玲玲 沈水仙 支涤静 陆 炜 陆 忠程若倩奚 立叶 蓉 赵诸慧 郑章乾 章淼滢 罗飞宏
生长分化因子-5(GDF5)基因由Hotten 等[1]于1994 年从人的cDNA 库中首次克隆得到。GDF5 基因的表达主要集中于胚胎软骨间充质细胞聚集区、长骨发育的软骨核以及即将形成关节的区域[2],在胚胎发育,调节细胞分化,促进肢体发育,修复肌腱、韧带、皮肤、神经、骨和软骨损伤等方面起到重要的作用[3~5]。GDF5 基因敲除的小鼠,其骨骼和关节韧带发育受损,表现出四肢短缩和体重减轻的表型[6,7]。
GDF5 作为软骨形成、骨骼发育的重要调节因子,近年受到广泛的关注与研究。国外多篇文献报道了GDF5 单核苷酸多态性(SNP)与身高的相关性。然而,迄今为止尚未见中国人群GDF5 基因多态性与身高相关性的报道,鉴于以上研究背景,本研究选择GDF5 基因rs224331 位点,分析其多态性与中国汉族青少年身高、体重和BMI 的关联性。
1 方法
1.1 纳入和排除标准 ①2007 年某技术学校入学新生,年龄和民族不限;②身体发育成熟,处于Tanner Ⅴ期;③排除先天发育畸形或后天器质性畸形如骨折等;④排除甲状腺疾病、垂体疾病、肿瘤和各种代谢性疾病。
1.2 研究现场 2007 年9 月20 ~22 日,以江苏省泰州市某技术学校新生入学体检为契机,在保证完成正常体检项目的前提下,本研究课题组儿科内分泌专科医生进行身高和体重测量,并对本研究纳入标准中的身体发育成熟期进行判断,对排除标准中的疾病状态进行排除。并于正常体检肝功能检测需要采血时征得体检学生同意仅进行GDF5基因检查。
1.3 标本采集 ①身高测量:被测者赤脚站立,立正姿势站在身高计的底板上,上肢自然下垂,足跟并拢,足尖分开约呈60 度角,双眼平视前方,脚跟、骶骨部及两肩胛骨(3点)靠紧身高计的立柱。测量者站在被测量者的一侧,移动身高计的水平板至被测量者的头顶,使其松紧度适当,记下读数,精确至0.1 cm。②体重测量:被测量者赤足,男生着短裤,女生着短裤、短袖衫,站在秤台中央,测试读数以kg 为单位,精确至0.1 kg。③抽取静脉血2 mL,EDTA 抗凝,-20℃冰箱冻存。身高、体重和静脉血均于体检日上午空腹状态下采集。
1.4 超重和肥胖标准 根据2008 年WHO 推荐的成人超重及肥胖标准[8]:体重正常:18.5≤BMI <25,超重:25≤BMI <30,肥胖:BMI≥30。
1.5 基因组DNA 提取 采用TIAGEN 血液基因组DNA提取试剂盒。A260/A280为1.6 ~2.0,并将样本浓度稀释至15 ~20 ng·μL-1。
1.6 SNP 位点选择 rs224331 是位于GDF5 基因外显子区域的一个错义突变,引起Ser276Ala 氨基酸改变。该位点与GDF5 基因内的多个位点(如rs143383、rs143384 和rs224333 等)连锁,r2分别为0.933、0.967 和0.967。图1所示的LD Plot 图显示了该基因各个SNP 位点之间的连锁情况,方块颜色的差异表示连锁程度不同,颜色越深表示连锁程度越高。

图1 GDF5 基因多态性位点连锁分析Fig 1Linkage disequilibrium analysis of GDF5 gene polymorphism loci
1.7 SNP 位点分型 采用ABI 公司Taqman 探针,C__25619958_10 。反应总体积为5 μL:包括40 ×探针0.125 μL,2 × Mix 2. 5 μL,DNA 1 μL,ddH2O 1. 375 μL。采用Roche LightCycler 480 PCR 仪进行扩增反应,PCR 反应条件如下:95℃预变性10 min;92℃变性15 s,60℃延伸1 min,重复45 个循环;60℃30 s,40℃1 s。rs224331 位点分型结果如图2 所示。

图2 rs224331 位点分型散点图Fig 2 Endpoint fluorescence scatter plot of rs224331
1.8 统计学分析 应用Execl 软件建立数据库,采用SPSS15.0 软件进行统计分析。计量资料以±s 表示。身高、体重和BMI 行正态分布检验,不同基因型间比较正态分布采用单因素方差分析,偏态分布采用非参数秩和检验。基因型及其等位基因频率分布进行Hardy-Weinberg 平衡检验。基因频率比较采用χ2检验。
2 结果
2.1 研究人群的一般情况 应当参与体检的新生共1 790名,年龄17 ~22 岁,均为汉族,青春发育均达到Tanner Ⅴ期。97.9%(1 754/1 790)SNP 分型成功,男生859 名,年龄(19.5 ±0.9)岁,身高(172.3 ±5.7)cm,体重(63.0 ±10.5)kg,BMI (21.2 ±3.3)kg·m-2,达超重标准76 名(8.8%),达肥胖标准21 名(2.4%);女生895 名,年龄(19.2 ±0.9)岁,身高(160. 0 ± 5. 3)cm,体重(52. 2 ± 6. 9)kg,BMI(20.4 ±2.5)kg·m-2,达超重标准37 名(4.1%),达肥胖标准2 名(0.2%)。
2. 2 连锁不平衡检验 表1 显示,rs224331 位点最常见的基因型是AA(51.7%),其次是AC(39.6%),CC 基因型最为少见(8.7%)。在Hardy-Weinberg 平衡检验中,无论是总样本还是男生、女生,rs224331 位点的基因型和等位基因频率均符合Hardy-Weinberg 平衡。
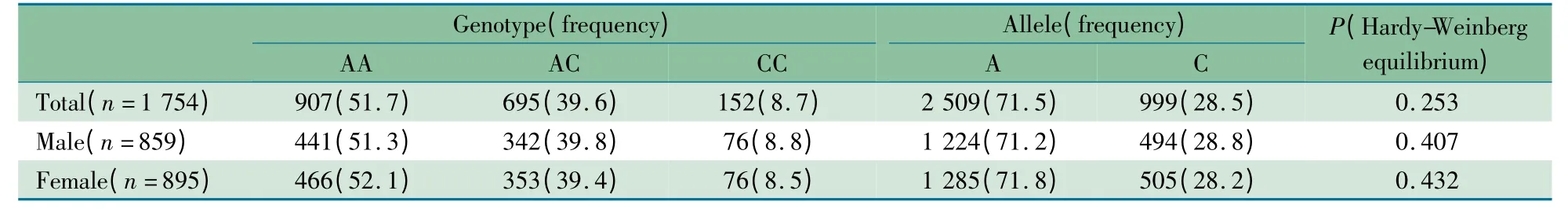
表1 GDF5 基因rs224331 位点基因型和等位基因频率分布[n(%)]Tab 1 Genotype and allele frequencies of the SNP (rs224331)of the GDF5 gene in Chinese adults[n(%)]
2.3 GDF5 基因rs224331 多态性与身高、体重和BMI 的相关性 ①与身高的相关性(表2):男生和女生的身高均呈正态分布,单因素方差分析结果显示,男生和女生rs224331位点的基因型分布均与身高无相关性,P 分别为0.728 和0.723。②与体重、BMI 的相关性(表2):男生和女生的体重、BMI 均呈偏态分布,非参数秩和检验结果显示,男生rs224331 位点不同基因型分布和体重、BMI 存在相关性,P分别为2.32 ×10-4和3.19 ×10-4;女生无显著相关性,P分别为0.713 和0.921。
GDF5 基因rs224331 位点不同基因型与身高、体重和BMI 线性回归分析显示(表3),AC 基因型的男生体重和BMI 均高于AA 基因型,P 均为0.002;女生则无此差异。
表2 GDF5 基因rs224331 位点不同基因型身高、体重和BMI 比较(±s)Tab 2 Relationship between rs224331 polymorphism and height,weight and BMI(±s)
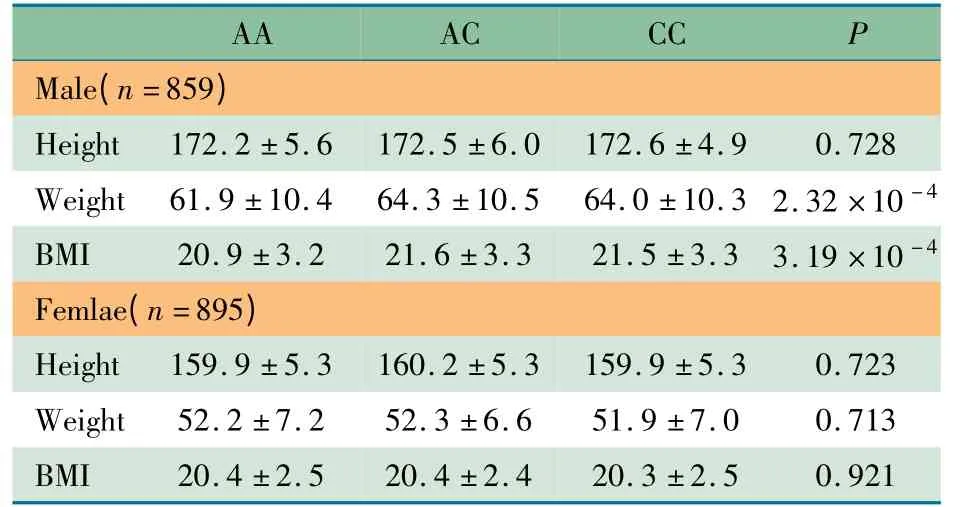
表2 GDF5 基因rs224331 位点不同基因型身高、体重和BMI 比较(±s)Tab 2 Relationship between rs224331 polymorphism and height,weight and BMI(±s)
Notes The unit of height,weight and BMI was cm,kg and kg·m -2,respectively
AA AC CC P Male(n=859)0.728 Weight 61.9 ±10.4 64.3 ±10.5 64.0 ±10.3 2.32 ×10 -4 Height 172.2 ±5.6 172.5 ±6.0 172.6 ±4.9 BMI 20.9 ±3.2 21.6 ±3.3 21.5 ±3.3 3.19 ×10 -4 Femlae(n=895)0.723 Weight52.2 ±7.252.3 ±6.651.9 ±7.00.713 Height 159.9 ±5.3 160.2 ±5.3 159.9 ±5.3 0.921 BMI 20.4 ±2.5 20.4 ±2.4 20.3 ±2.5

表3 GDF5 基因rs224331 位点不同基因型对身高、体重和BMI影响的线性回归分析Tab 3 Linear regression analysis of the relationship between rs224331 polymorphism and height,weight and BMI
3 讨论
国外在GDF5 基因多态性与软骨形成、骨骼发育和身高的相关性研究中,多选择rs143384 和rs143383 位点,一般认为当两个位点r2>0.8,则其中一个位点可代替另一个位点[9]。图1 为中国汉族人群的GDF5 基因多态性位点连锁关联,本研究中选取的rs224331 位点与rs224333,rs224330,rs143383,rs143384 位点的r2均>0.8,说明这些位点间的连锁性强。
本研究为了探讨中国汉族人群中GDF5 基因多态性与身高的相关性,选取了引起错义突变的一个常见位点rs224331,发现GDF5 基因多态性位点rs224331 不同基因型分布与身高之间无明显的相关性,与国外多项研究结论相反。芬兰6 114 人的GWAS 研究发现[10],GDF5 基因多态性位点rs6060369 与身高相关(P =9.7 ×10-7);荷兰鹿特丹市的人群研究发现[11],GDF5 基因rs143383 多态性与女性身高存在关联性;8 184 名欧洲裔美国儿童中验证了之前文献报道的与身高相关性最强的51 个位点的SNP[12],结果发现其中16 个位点在该人群中与身高相关,包括GDF5多态性位点rs4911494(P=1.63 ×10-3)。本文结果与上述文献结论不一致的原因可能为:①种族差异:rs224331 位点的等位基因分布频率可能在不同人种中存在差异,上述文献报道均为欧洲人种GDF5 基因多态性与身高有相关性,本研究人群为中国汉族人。②身高是人体生长发育重要的生理参数,由多种因素共同作用[13],虽然遗传因素起到80%的作用,但营养、疾病和环境等因素的作用亦不能忽视[14,15]。③本研究人群样本量可能影响结果的准确性。④本研究仅选取了rs224331 位点,尽管与文献[10~12]选取的基因位点有很强的关联性,但对更加全面的诠释GDF5与身高的关联性可能产生影响。
虽然本研究没有得到与国际相对一致的GDF5 基因多态性与身高之间相关性的结论,但却发现GDF5 基因rs224331 位点多态性与男生体重、BMI 存在相关性。与男生不同的是,本研究在女生中并未发现GDF5 基因多态性与体重、BMI 的相关性。分析其原因:本课题组对上海15万儿童青少年肥胖的患病率调查发现,6 ~10 岁为患病率增高期,10 ~18 岁为逐渐降低期,女性的肥胖患病率显著低于男性[16];与西方国家的肥胖患病率随年龄增加而升高的现象有着本质的不同。本次研究与文献[16]一致,超重和肥胖检出率女生分别约为男生的50%和10%,因此,除遗传因素外,中国这一年龄段的女生更注重对体型的要求可能在更大程度上影响着体重的变化。其次,GDF5 基因可能与性别相关激素有关。例如转化生长因子-β(TGF-β)超家族的骨形成发生蛋白2(BMP2)与雌激素的作用相关,雌激素能够促进BMP2 的表达。因GDF5 与BMP2 结构和功能的相似性,推测其与性别相关激素可能有相关性,从而导致男生和女生在GDF5 基因多态性和体重、BMI 的相关性上出现差异[17]。
GDF5 基因rs224331 位点多态性与男生体重、BMI 相关,AC 基因型的男生体重和BMI 高于AA 基因型,AA 和CC 基因型,CC 和AC 基因型间差异无统计学意义。
总之,GDF5 基因多态性与身高、体重和BMI 的相关性结论不一。除了受研究对象种族、地域差异及其他环境因素的影响之外,不能排除其他已知的或未知的基因的协同或干扰作用。
[1]Hotten G,Neidhardt H,Jacobowsky B,et al. Cloning and expression of recombinant human growth/differentiation factor 5.Biochem Biophys Res Commun,1994,204(2):646-652
[2]Francis-West PH,Abdelfattah A,Chen P,et al. Mechanisms of GDF-5 action during skeletal development. Development,1999,126(6):1305-1315
[3]Hatakeyama Y,Tuan RS,Shum L. Distinct functions of BMP4 and GDF5 in the regulation of chondrogenesis. J Cell Biochem,2004,91(6):1204-1217
[4]Takahara M,Harada M,Guan D,et al. Developmental failure of phalanges in the absence of growth/differentiation factor 5.Bone,2004,35(5):1069-1076
[5]Oshin AO, Caporali E, Byron CR, et al. Phenotypic maintenance of articular chondrocytes in vitro requires BMP activity. Vet Comp Orthop Traumatol,2007,20(3):185-191
[6]Suto J. Identification of mutation in the growth differentiation factor 5 (Gdf5)gene in NC-brp/brp mice. J Vet Med Sci,2006,68(10):1121-1124
[7]Daans M,Luyten FP,Lories RJ. GDF5 deficiency in mice is associated with instability-driven joint damage, gait and subchondral bone changes. Ann Rheum Dis,2011,70(1):208-213
[8]http://www. who. int/dietphysicalactivity/childhood_what/en/index.html
[9]Wall JD,Pritchard JK. Haplotype blocks and linkage disequilibrium in the human genome.Nat Rev Genet,2003,4(8):587-597
[10]Sanna S,Jackson AU,Nagaraja R,et al. Common variants in the GDF5-UQCC region are associated with variation in human height. Nat Genet,2008,40(2):198-203
[11]Vaes RB,Rivadeneira F,Kerkhof JM,et al. Genetic variation in the GDF5 region is associated with osteoarthritis,height,hip axis length and fracture risk:the Rotterdam study. Ann Rheum Dis,2009,68(11):1754-1760
[12]Zhao J,Li M,Bradfield JP,et al. The role of height-associated loci identified in genome wide association studies in the determination of pediatric stature. BMC Med Genet,2010,11:96
[13]Lettre G.Genetic regulation of adult stature.Curr Opin Pediatr,2009,21(4):515-522
[14]Silventoinen K,Sammalisto S,Perola M,et al. Heritability of adult body height:a comparative study of twin cohorts in eight countries. Twin Res,2003. 6(5):399-408
[15]Perola M,Sammalisto S,Hiekkalinna T,et al. Combined genome scans for body stature in 6,602 European twins:evidence for common Caucasian loci. PLoS Genet,2007,3(6):e97
[16]Zhang M,Guo F,Tu Y,et al. Further increase of obesity prevalence in Chinese children and adolescents -cross-sectional data of two consecutive samples from the city of Shanghai from 2003 to 2008. Pediatr Diabetes,2012,6;9999(9999). Epub ahead of print
[17]Zhou S,Turgeman G,Harris SE,et al. Estrogens activate bone morphogenetic protein-2 gene transcription in mouse mesenchymal stem cells. Mol Endocrinol,2003,17(1):56-66
