水杨酸盐诱发大鼠听皮层中Egr-1基因表达的改变△
2012-12-04胡守森黄治物吴皓梅玲陈建勇
胡守森 黄治物,2 吴皓,2 梅玲,2 陈建勇,2
1 上海交通大学医学院耳科学研究所听觉科学实验室(上海200092);2 上海交通大学医学院附属新华医院耳鼻咽喉-头颈外科
水杨酸盐是一种在临床上广泛应用的解热镇痛抗炎类药物,长期使用会导致可逆性听力下降和耳鸣。动物实验中常利用水杨酸盐的耳毒性以及噪声暴露来进行耳鸣动物造模。动物行为学实验已经证实注射水杨酸盐后,动物可以产生耳鸣,在前期研究中给动物注射水杨酸盐后发现,急性注射可以使豚鼠的耳蜗神经活动平均功率谱(average spectrum of electrophysiological cochleoneural activity,ASECA)1kHz谱峰值和畸变产物耳声发射(DPOAE)幅值下降,而慢性注射则使两者的幅值上升[1,2]。研究发现急性注射水杨酸盐对耳蜗外毛细胞(OHC)外侧膜上的prestin蛋白表达无影响,而长期注射水杨酸盐后OHC膜上的prestin mRNA及蛋白表达呈可逆性上调,推测这可能是引起DPOAE幅值增加的主要原因[3]。以上研究表明,水杨酸盐致耳鸣极可能源自于耳蜗水平的结构功能改变向更高水平及中枢传递。由于听觉中枢存在一定的可塑性,那么源于外周的耳鸣长期持续对听觉中枢的不良刺激,听觉中枢是否会因此产生结构和功能方面的重组以及耳鸣产生的中枢具体机制尚不清楚。神经元的持续反应会导致某些基因表达的改变和新蛋白的合成,在噪声暴露的耳鸣动物模型中,观察到听皮层神经元的调谐频率降低,皮层同步化增加[4];有研究表明,早期生长反应基因-1(early growth response gene-1,Egr-1)在细胞外刺激和胞内基因表达改变中起重要的耦合作用,并调控下游长期反应基因(long term response genes)的表达,在神经元长时程可塑性中起着重要作用。Egr-1转录因子的激活依赖于细胞内钙离子浓度升高,通过胞内信号级联通路的放大,导致了一系列胞内目标蛋白的改变,将短时程信号改变转变成为Egr-1转录因子所作用的一系列长时程转变[5]。为此,本研究拟通过长期肌肉注射水杨酸盐建立大鼠耳鸣动物模型,观察建模过程中大鼠听皮层Egr-1的表达变化,以了解Egr-1是否参与了耳鸣的产生以及在耳鸣持续发展的过程中是否存在中枢可塑性,探讨耳鸣产生的中枢可塑性机制。
1 材料与方法
1.1 实验材料 水杨酸盐购自美国Sigma公司,以生理盐水溶解,其终浓度为100g/L;Trizol、PrimeScript®RT Master Mix(Perfect Real Time)试剂盒和 SYBR®Premix Ex TaqTM(Tli RNaseH Plus)试剂盒均购自日本Takara公司;Egr-1单克隆抗体购自美国CST公司;山羊抗兔IgG二抗购自北京鼎国生物技术公司;ECL(enhanced chemiluminescence)化学发光试剂盒购自美国 Millipore公司。
1.2 实验方法
1.2.1 实验动物分组及处理 8周龄SPF(specific pathogen free)级健康雄性SD大鼠(上海交通大学附属新华医院动物实验中心提供)84只,体重180~200g,耳廓反射阳性,无强噪声暴露及耳毒性药物使用史。随机分为7组,每组12只,分别为:正常对照组(A组);急性注射2小时组(B组,400mg/kg水杨酸盐肌肉注射,2.5小时后断头);慢性注射3天组(C组,200mg/kg水杨酸盐肌肉注射,2次/天,间隔8小时,连续注射3天后在第4天断头);慢性注射7天组(D组,200mg/kg水杨酸盐肌肉注射,2次/天,间隔8小时,连续注射7天后在第8天断头);慢性注射14天组(E组,200mg/kg水杨酸盐肌肉注射,2次/天,间隔8小时,连续注射14天后在第15天断头);停药后恢复14天组(F组,慢性水杨酸盐注射14天后再停止注射恢复14天断头);停药后恢复28天组(G组,慢性水杨酸盐注射14天后再停止注射恢复28天断头)。各组大鼠断头处死后迅速取听皮层,其中6只用于检测Egr-1基因的变化,另外6只用于检测Egr-1蛋白的改变。
1.2.2 SYBR Green实时荧光定量PCR检测各组大鼠听皮层Egr-1mRNA的表达 所有引物根据Genbank的登录号,查找mRNA序列,由日本TaKaRa公司设计,由上海生工生物工程技术服务有限公司合成。引物序列如下:GAPDH,GAPDH-F:GGCACAGTCAAGGCTGAGAATG GAPDH-R:ATGGTGGTGAAGACGCCAGTA Egr-1,Egr-1-F:GAACAACCCTACGAGCACCTG Egr-1-R:GCCACAAAGTGTTGCCACTG各组大鼠取听皮层后,立即放入液氮中,随后转入-80℃冰箱保存。样本收集完后用Trizol法分别抽提总RNA。紫外分光光度计检测其在260nm、280nm的吸光度值,计算 OD 260/OD280的比值鉴定RNA的纯度。
采用TaKaRa公司的PrimeScript®RT Master Mix(Perfect Real Time)试剂盒进行cDNA的合成,按照说明书配制反应液,反转录程序:37℃15 min;85℃5sec。将cDNA溶液置于-20℃保存备用。取逆转录产物40倍稀释,real-time PCR反应程序按照TaKaRa公司的SYBR®Premix Ex TaqTM(Tli RNaseH Plus)试剂盒说明书进行设置:Stage 1:预变性95℃30秒,1Cycle;Stage 2:PCR扩增反应95℃5秒,60℃34秒 ,40Cycles;Stage 3:95℃15秒,65℃60秒 ,95℃15秒,1Cycle。
1.2.3 Western blot分析Egr-1蛋白的表达 大鼠听皮层超声粉碎(3秒2次,间隔2秒)后直接加入PMSF和蛋白裂解液。加入700μl组织裂解液,离心并取上清液。各组取总蛋白80μg,经10%SDS-PAGE凝胶电泳分离,后转移至PVDF膜,然后用封闭液封闭1小时,加入一抗4℃过夜。第二天,复温半小时后用TBST洗3次,加入二抗孵育1.5小时,TBST洗三次,最后在暗室内加入ECL化学发光底物曝光。选用β-Actin作为内参照,并用图像分析系统分析结果。
1.3 统计学处理 采用SPSS 11.5软件进行统计学处理,组间均数差异性比较采用单因素方差分析,P<0.05为差异有统计学意义,其中实时荧光定量PCR数据收集主要由The Applied Biosystems 7500Real Time PCR System自带软件完成。各组基因表达变化采用比较阈值法即2-ΔΔCt法计算。
2 结果
2.1 各组大鼠听皮层中Egr-1mRNA的表达各组大鼠听皮层中Egr-1mRNA表达水平随着水杨酸盐注射次数的增加呈逐渐下降趋势,慢性注射3、7、14天组(C、D、E组)与正常对照组、急性注射2小时组、停药后恢复14天组、停药后恢复28天组(A、B、F、G组)相比,Egr-1mRNA 表达水平均明显下降,差异具有统计学意义(P<0.05);正常对照组、急性注射2小时组、停药后恢复14天组、停药后恢复28天组的差异无统计学意义(P>0.05)(表1,图1)。
表1 各组大鼠听皮层Egr-1mRNA表达水平的比较(±s)

表1 各组大鼠听皮层Egr-1mRNA表达水平的比较(±s)
注:Ct表示循环数;ΔCt=Egr-1Ct的平均值-GAPDH Ct的平均值;ΔΔCt= 各组的ΔCt-正常对照组的ΔCt;*与正常对照组比较,P<0.05
组 别 Egr-1Ct GAPDH Ct ΔCt ΔΔCt 2-ΔΔCt正常对照组 26.53±0.15 25.44±0.61 1.10±0.53 0.00±0.53 1.85±0.33.05±0.34急性注射2小时组 27.11±0.15 25.52±0.42 1.60±0.53 0.50±0.53 0.74±0.24慢性注射3天组 26.61±0.98 24.70±1.43 1.91±0.55 0.81±0.55 0.61±0.25*慢性注射7天组 26.47±0.70 24.61±0.83 1.85±0.41 0.76±0.41 0.61±0.22*慢性注射14天组 28.35±0.82 26.07±0.44 2.28±0.77 1.19±0.77 0.49±0.23*停药后恢复14天组 26.81±0.61 25.18±0.59 1.63±0.78 0.54±0.78 0.78±0.46停药后恢复28天组 26.94±0.65 25.51±1.07 1.42±0.53 0.33±0.53 0

图1 实时荧光定量PCR检验各组中Egr-1mRNA表达的变化
2.2 各组大鼠听皮层Egr-1蛋白的表达 Western blot印迹结果显示,各组大鼠听皮层均有Egr-1蛋白表达,慢性注射3天组、慢性注射7天组、慢性注射14天组(C、D、E组)Egr-1蛋白表达降低(P<0.05),其余各组蛋白表达差异无统计学意义(P>0.05)(图2、3)。
3 讨论
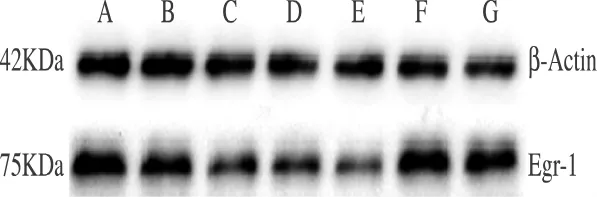
图2 Western-blot检测各组中Egr-1蛋白表达的变化
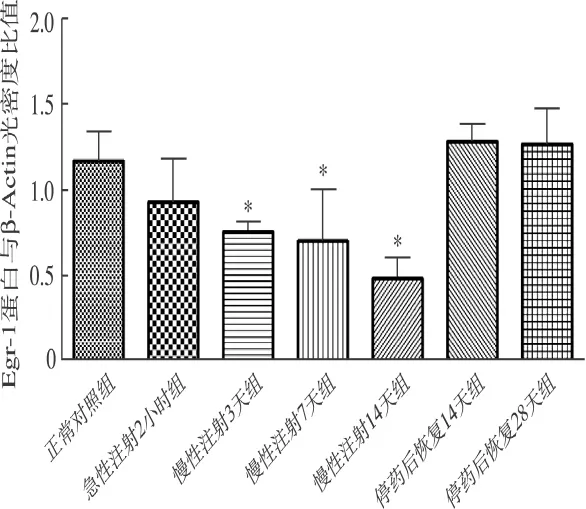
图3 各组大鼠听皮层Egr-1蛋白与β-Actin的光密度比值
耳蜗外毛细胞(OHC)是水杨酸盐耳毒性作用的靶细胞,外毛细胞的电能动性在听觉灵敏性和特异性方面起着至关重要的作用。研究观察了反应OHC电能动性的DPOAE幅值和表达在OHC侧膜上的驱动蛋白prestin的表达量,证实了水杨酸盐所致耳鸣极可能源自于耳蜗水平[1~3]。然而耳鸣的产生并不仅仅只是停留在耳蜗水平,而是涉及到听觉通路的更高层面,尤其是耳鸣是否存在中枢化现象,耳鸣的中枢机制是否涉及到中枢化问题,以及是否会诱发多种神经递质和即刻早期基因的改变[6],目前尚无确切的实验依据。
Egr-1在神经元活动中维持着可塑性的改变。Egr-1是一种可被不同形式神经活性迅速诱导的即刻早期基因,它介导了细胞的增殖与分化,参与细胞的凋亡过程和组织的损伤修复,尤其在维持神经元可塑性改变方面也发挥着重要的作用[1]。已有研究证实突触可塑性常表现为长时程增强(longterm potentiation,LTP),而Egr-1的表达能作为引发LTP的分子标识(LTP常常伴随着Egr-1表达的升高)[8],用于反应LTP后的神经元出现结构重组的情况,并且在大脑皮质结构的刺激依赖性生长中起着重要的作用[7]。Nunes等[9]的研究观察到小鼠暴露在丰富行为学习环境中一个月,伴随着LTP的增强,扣带回皮质前端的Egr-1表达升高;给成年大鼠做运动训练,大鼠脊髓神经元中Egr-1的表达也升高。有研究发现,在海马和杏仁体中Egr-1在长时程增强和学习过程中表达上调[10],而在Egr-1敲除小鼠中观察到晚时相声音恐惧记忆受损,但短时的痕迹和场景记忆却不受影响,表明在晚时相听觉恐惧记忆中Egr-1极可能发挥关键性作用[10]。这些都说明Egr-1与神经可塑性,特别是表现为长时程增强的突触可塑性机制密切相关[11]。
Yu等[12]对豚鼠进行连续2周的水杨酸盐注射后,发现动物的DPOAE幅值逐渐上升,prestin表达上调,且耳蜗中的Egr-1表达水平较对照组上升了4.71倍;免疫荧光显示Egr-1表达在耳蜗外毛细胞核,表明Egr-1可能参与了prestin表达的上调。Ko等[10]发现用低强度的短阵脉冲刺激(theta burst stimulation,TBS)正常小鼠听皮层后可引发持续40分钟的突触反应的长时程增强,而在Egr-1基因敲除小鼠,用低强度的短阵脉冲刺激听觉皮层所引起的LTP被明显减弱或完全阻断。这些结果有力的证明了Egr-1有利于听皮层区域的LTP和听皮层的可塑性形成,进一步表明Egr-1与听觉中枢神经元可塑性有着密切联系。
从本研究结果看,急性注射水杨酸盐大鼠听皮层的Egr-1表达略有下降,但与正常对照组相比无明显差异,而慢性注射水杨酸盐后大鼠听皮层Egr-1表达下降,与对照组比较差异有统计学意义(P<0.05),与在耳蜗外毛细胞外侧膜上的prestin蛋白在急性和慢性注射水杨酸盐时表达上升的结果(不同时相的变化规律)[9]相比较,相关性极好。大鼠的LTP按其时程分为3种:LTP1持续大概2个小时,LTP2持续大概4天,LTP3持续大概23天。第一种叫E-LTP,后两种统称L-LTP;研究表明Egr-1在L-LTP的稳定中发挥着重要的作用[9],且L-LTP(而非E-LTP)要求有基因转录和蛋白合成,因此,本研究中慢性注射水杨酸盐后大鼠听皮层Egr-1的表达下降,推测其导致了听皮层突触反应长时程增强,表明听皮层发生了神经可塑性的改变。
长期注射水杨酸盐后,DPOAE幅值增加,OHC膜上的prestin蛋白表达上调,造成了耳蜗外毛细胞电能动性的增强,引起ASECA的1kHz谱峰值升高,说明耳蜗神经活动增强[1~3],进一步上传使听皮层活动性升高,感知为耳鸣。这种长期活动过度的刺激,导致机体为了维持自身稳态平衡使神经元活动恢复到正常范围,启动了一系列补偿措施,其中之一就是Egr-1的表达下降;而Egr-1表达降低将不利于LTP,使神经元产生适应不良的可塑性。
水杨酸盐的耳毒性作用具有剂量依赖性,即用药剂量越大、用药时间越长,其所导致的耳蜗损伤越大,这与本研究观察到的Egr-1在慢性注射水杨酸盐3天、7天和14天组的耳鸣大鼠听皮层表达呈下降趋势相一致,也与早期研究中慢性注射水杨酸盐后,豚鼠的DPOAE和ASECA逐渐升高在时相上极其相似[1,2]。水杨酸盐对耳蜗的损伤也具有可逆性,停止注射水杨酸盐后DPOAE、ASECA幅值和prestin蛋白表达可以恢复正常[3]。本研究结果显示停药后恢复14天组、停药后恢复28天组大鼠听皮层Egr-1的表达与正常对照组比较差异无统计学意义,说明听皮层中Egr-1的表达改变是可逆性的(恢复正常)。此外,Egr-1也能通过辅助抑制因子(NGFI-A binding 2,NAB2)产生自身表达的负反馈调节来维持表达的平衡从而使表达恢复正常。
总之,水杨酸盐所致耳鸣不仅仅发生在耳蜗水平,更涉及到高级听觉中枢。长期注射水杨酸盐后,大鼠听皮层Egr-1表达的改变证实了Egr-1参与了听皮层可塑性变化。然而听觉系统的可塑性涉及到听觉通路的多个层面,除听皮层外,Egr-1在耳蜗核、下丘的表达如何还有待进一步的研究。
1 Cazals Y,Horner KC,Huang ZW.Alterations in average spectrum of cochleoneural activity by long-term salicylate treatment in the guinea pig:aplausible index of tinnitus[J].J Neurophysiol,1998,80:2 113.
2 Huang ZW,Luo Y,Wu Z,et al.Paradoxical enhancement of active cochlear mechanics in long-term administration of salicylate[J].J Neurophysiol,2005,93:2 053.
3 Yang K,Huang ZW,Liu ZQ,et al.Long-term administration of salicylate enhances prestin expression in rat cochlea[J].Int J Audiol,2009,48:18.
4 Engineer ND,Riley JR,Seale JD,et al.Reversing pathological neural activity using targeted plasticity[J].Nature,2011,470:101.
5 Thiel G,Mayer SI,Muller I,et al.Egr-1-A Ca2+-regulated transcription factor[J].Cell Calcium,2010,47:397.
6 Browne CJ,Morley JW,Parsons CH.Tracking the expression of excitatory and inhibitory neurotransmission-related proteins and neuroplasticity markers after noise induced hearing loss[J].PLoS One,2012,7:e33272.
7 Platenik J,Kuramoto N,Yoneda Y.Molecular mechanisms associated with long-term consolidation of the NMDA signals[J].Life Sci,2000,67:335.
8 Shum FW,Wu L J,Zhao MG,et al.Alteration of cingulate long-term plasticity and behavioral sensitization to inflammation by environmental enrichment[J].Learn Mem,2007,14:304.
9 Nunes AC,Duarte RB,Sousa TB,et al.Expression of the immediate-early gene Egr-1and substance P in the spinal cord following locomotor training in adult rats[J].Brain Res,2010,1 345:125.
10 Ko SW,Ao HS,Mendel AG,et al.Transcription factor Egr-1is required for long-term fear memory and anxiety[J].Sheng Li Xue Bao,2005,57:421.
11 Knapska E,Kaczmarek L.A gene for neuronal plasticity in the mammalian brain:Zif268/Egr-1/NGFI-A/Krox-24/TIS8/ZENK?[J].Prog Neurobiol,2004,74:183.
12 Yu N,Zhu ML,Johnson B,et al.Prestin up-regulation in chronic salicylate(aspirin)administration:an implication of functional dependence of prestin expression[J].Cell Mol Life Sci,2008,65:2 407.
