孟氏隐唇瓢虫成虫自残幼虫的研究
2012-11-22蒋瑞鑫张宇宏吴红胜谢佳沁
蒋瑞鑫,张宇宏,吴红胜,刘 平,谢佳沁,庞 虹
(有害生物控制与资源利用国家重点实验室/中山大学昆虫学研究所,广州510275)
自然种群中,自残行为是一种正常的种群调控机制 (Fox,1975)。昆虫的自残习性相当严重(王孟卿和彩万志,2004)。就捕食性瓢虫而言,自残行为普遍存在,已经被认为是正常觅食行为的一部分 (Dixon,2000)。因为瓢虫初孵幼虫通过取食同种卵能提高自身的存活率 (Gagnéet al.,2002),而且幼虫之间的自残使其可以顺利化蛹羽化 (王甦等,2010),所以自残对于提高瓢虫在逆境中的生存率具有适应意义 (Agarwala and Dixon,1992)。但是目前瓢虫自残的研究多集中在食蚜瓢虫,食蚧瓢虫的研究和评论很少,这可能与食蚧瓢虫觅食猎物的方式相关,它们的卵和低龄幼虫经常藏于它们的猎物之下,因此降低了与高龄幼虫相遇的风险 (Dixon,2000)。在田间,同种幼虫大量存在于猎物斑块时,其成虫就会离开该斑块而寻找新的猎物,所以成虫残杀幼虫的现象也就相对不普遍,所以目前瓢虫的自残研究都集中在卵和幼虫阶段。但是在室内大量扩繁捕食性瓢虫时,成虫和幼虫常常会共存很长一段时间,所以了解成虫对幼虫的自残习性有助于改进人工饲养技术,提高产量。
孟氏隐唇瓢虫Cryptolaemus montrouzieri Mulsant原产于澳大利亚,经过120多年的引种扩散,目前已在世界范围内的20多个国家和地区定殖 (蒋瑞鑫等,2009)。为了能商品化生产该瓢虫,不少学者对其人工饲料和保种技术进行了深入地研究(李丽英,1993;庞虹等,1996;Venkatesan et al.,2001),但是对于该瓢虫的自残行为没有相应的报道。而对于瓢虫而言,非同血缘间的自残往往要比同血缘间的自残更为严重 (Osawa,1989;王甦等,2010),所以从生产的角度出发,本文就孟氏隐唇瓢虫成虫自残非同血缘幼虫的行为开展研究,以提高大规模人工饲养孟氏隐唇瓢虫的产量。
1 材料与方法
1.1 昆虫饲养
试验所用孟氏隐唇瓢虫C.montrouzieri于2006年引种自印度,在中山大学昆虫园室内以生活在南瓜果实上的双条拂粉蚧Ferrisia virgata(Cockerell)和柑桔粉蚧Planococcus citri(Risso)为食,在玻璃铝合金纱网的养虫笼 (45.0 cm×36.0 cm×33.0 cm,铝合金及120目塑料纤维纱网制)中连续饲养3年以上。所有供试昆虫的饲养环境为T=25±1℃,RH=50±10%,L∶D=14∶10。
1.2 成虫对高龄幼虫的自残行为
在不同的养虫笼中分别挑取4龄幼虫1头(蜕皮﹤48 h)和成虫1头 (羽化<14 d)置于小塑料培养皿中,不提供任何食物,分别于12 h、24 h、36 h和48 h记录幼虫的存活情况。该试验设置雄成虫和雌成虫2个处理,每个处理20个样本。计算每个处理中20个幼虫样本在每个时间点的累积死亡率。累积死亡率=本时间点之前 (包括本时间点)发生自残死亡的总样本数/样本总数(20)。试验中所用昆虫均为非同血缘关系,如无特别说明,下同。
1.3 成虫对低龄幼虫的自残行为
取孟氏隐唇瓢虫雌虫1头 (羽化10~30 d,并已交配开始产卵)置于小塑料培养皿 (5.5 cm×1.5 cm)中,饥饿24 h后,在养虫笼中挑取2龄孟氏隐唇瓢虫幼虫1头,放入上述载有雌虫的培养皿中,观察雌虫对幼虫的自残过程,并记录在整个过程中孟氏隐唇瓢虫雌虫各种行为 (如表1)发生的次数和相应持续时间,以及成虫攻击幼虫的部位。试验设置两个处理:1)幼虫体被蜡丝,即正常幼虫;2)幼虫体表无蜡丝 (蜡丝用毛笔轻轻扫去),称之为裸虫。每个处理15个重复。自残过程:从雌虫第一次攻击幼虫开始直到幼虫被攻击致死的整个过程。幼虫死亡的确定:幼虫遭受雌虫攻击后,完全失去行动能力,则视为死亡。

表1 孟氏隐唇瓢虫成虫的各种行为定义Table 1 The definition of behaviors of Cryptolaemus montrouzieri adult
1.4 自残幼虫对成虫生殖力的影响
取在养虫笼中刚羽化的孟氏隐唇瓢虫,雌雄一一配对后放入小塑料培养皿 (6 cm×1.5 cm)中饲养,并从南瓜上刮取充足的粉蚧和粉蚧的卵鞘供其取食和产卵,每24 h更换一次饲料和卵鞘,并观察其是否交尾,交尾后每24 h收集为其提供产卵的粉蚧卵鞘,在显微镜下检查产卵数,取连续3 d都产卵,并且3 d的日平均产卵量在5粒以上 (包括5粒)的雌虫用于试验。试验环境:T=25±1℃,RH=50±10%,L∶D=14∶10。
将上述孟氏隐唇瓢虫雌雄成虫置于小塑料培养皿中,然后从养虫笼中挑取4龄孟氏隐唇瓢虫幼虫2头,用毛笔扫去体表白色蜡质,接着用手术剪剪除幼虫的6根足 (以防止幼虫移动)后作为食料让成虫取食12 h,然后把雌雄成虫转入到新的培养皿 (6 cm×1.5 cm),并在培养皿中放入一小团棉花作为雌虫的产卵基质让其产卵,12 h后检查雌虫的产卵量,然后将成虫再转入到新的培养皿用新鲜的4龄断足幼虫继续饲养,如此循环,持续10 d。15个重复。对照组的饲料为柑桔粉蚧。待瓢虫产卵后,在两种处理中分别随机挑取30粒,置于小塑料培养皿中,每皿1粒,于室温下培养,每隔12 h观察一次,记录卵的孵化时间和孵化数,按表2所列公式计算所需参数,如果卵超过10 d未孵化,则视为死亡。

表2 试验所得参数及其计算方法Table 2 The parameters of present research andthe related pathway of figurations
1.5 数据分析
对不同性别成虫自残幼虫48 h对应的累积死亡率进行费雪精确检验 (Fisher's exact test);以幼虫形态 (正常幼虫和裸虫)和饲料为独立变量分别对试验1.3和1.4中观测的参数进行独立T检验;所有数据分析在统计软件SPSS17.0中完成。
2 结果与分析
孟氏隐唇瓢虫在未饥饿的情况下,雌虫在48 h内自残4龄幼虫造成的累积死亡率为60%,而雄虫则为15% (图1),差异显著 (Fisher's exact test:P(2-sided)<0.01),表明雌虫对幼虫的残杀要比雄虫更为强烈。

图1 孟氏隐唇瓢虫成虫残杀4龄幼虫的累积死亡率Fig.1 The accumulative mortality of the 4th instar larvae of Cryptolaemus montrouzieri killed by adults

图2 孟氏隐唇瓢虫雌虫对2龄幼虫自残过程中的行为发生次数Fig.2 The happening behavior times in the cannibalism process of Cryptolaemus montrouzieri female to the 2nd instar larvae
由图2可知,孟氏隐唇瓢虫雌虫对不同形态2龄幼虫的自残过程中,均未观察到雌虫排泄,对于正常幼虫的自残所涉及的4种行为次数均显著高于裸虫处理 (t搜索=6.369,df=14.231,P(2-tailed)< 0.01;t梳理=6.177,df=14.080,P(2-tailed)<0.01;t攻击=7.89,df=14.883,P(2-tailed)< 0.01;t休息=2.215,df=18.096,P(2-tailed)=0.04)。此外,如表3所示,除了休息行为发生的总时间无显著差异外 (t=1.257,df=25.198,P(2-tailed)=0.22),雌虫对正常幼虫自残中在各行为上花费的总时间均显著高于裸虫处理 (t搜索=5.214,df=14.016,P(2-tailed)< 0.01;t梳理=4.449, df=14.006,P(2-tailed)< 0.01;t攻击=2.594,df=28,P(2-tailed)=0.015)。但是,雌虫对裸虫自残过程中,攻击行为的总时间占整个自残总时间的比例为98.21%(图3),显著高于正常幼虫处理中的41.76%(t= -11.783,df=15.105,P(2-tailed)<0.01),然而,孟氏隐唇瓢虫雌虫最后一次攻击正常幼虫和裸虫的平均时间分别为406.87 s和405.07 s,无显著差异 (t=0.014,df=28,P(2-tailed)=0.989),分别占到各自攻击总时间比例为70.55%(正常幼虫处理)和97.25%(裸虫处理),所以对雌虫而言,大约30%的攻击时间是用于咬击正常幼虫体表的蜡丝,而在裸虫处理中,成虫几乎只攻击幼虫一次,就可以把幼虫致死。
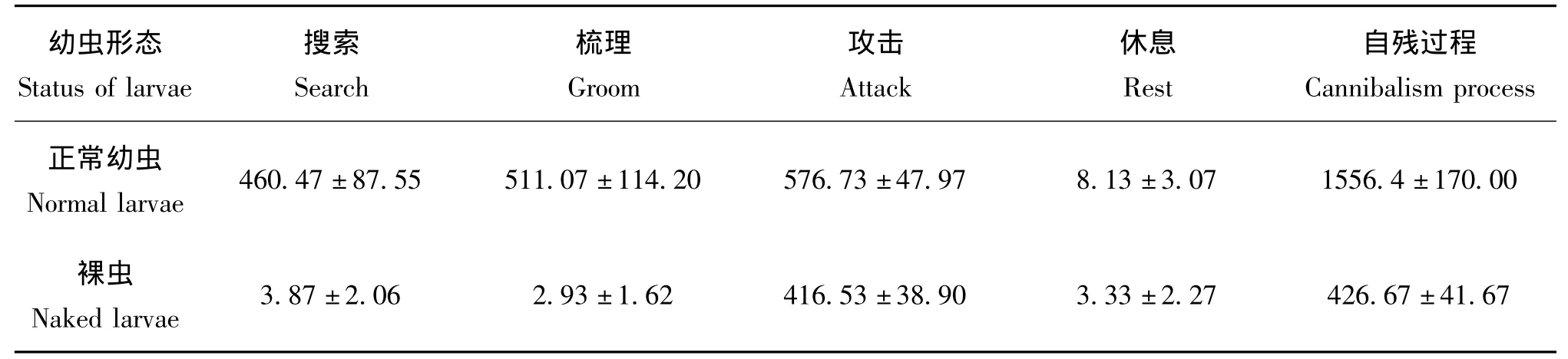
表3 孟氏隐唇瓢虫雌虫自残幼虫过程中各种行为的耗时 (单位:秒)Table3 ThetimespendinbehaviorsintheprocessofCryptolaemusmontrouzierifemalecannibalizelarvae(s)
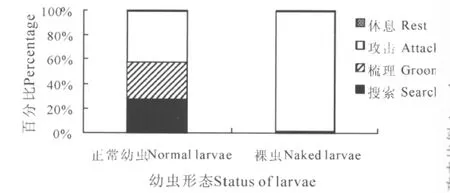
图3 孟氏隐唇瓢虫雌虫自残幼虫各行为占自残总耗时的比例Fig.3 ThetimepercentspendinbehaviorsintheprocessofCryptolaemusmontrouzierifemalecannibalizelarvae

图4 孟氏隐唇瓢虫幼虫不同部位受攻击的比例Fig.4 TheattackedpercentageofdifferentpartsofCryptolaemus montrouzierilarvae
自残过程中,雌虫对两种不同形态幼虫的头,胸,腹均发起攻击,除了头部外,不同处理相同部位受攻击比例差异不显著 (t头=2.153,df=27.899,P(2-tailed)=0.04;t胸= -0.61, df=17.617,P(2-tailed)=0.952;t腹= -0.865,df=20.461,P(2-tailed)=0.397)。但各部位受攻击比例在同一幼虫处理中差异显著 (Friedman检验:χ2裸虫=21.814,df=2,P <0.01;χ2正常幼虫=23.424,df=2,P<0.01),如图4所示,以腹部受攻击的比例最高,分别为 79.74% (正常幼虫)和 87.22%(裸虫),头部和胸部受到的攻击次数相对较少。
不同饲料饲养对孟氏隐唇瓢虫的日均产卵率影响显著 (t=-5.214,df=11.21,P(2-tailed)<0.01),取食粉蚧的孟氏隐唇瓢虫10d内日均产卵率为90%,而取食幼虫的仅为42.67%。如图5所示,取食粉蚧的雌虫日产卵率除第一天低于80%外,其它天数的产卵率都在80%以上,而取食幼虫的雌虫日产卵率前5d都在50%以下,随后逐渐增加,到第9d、第10d达到最大,并趋于相对平稳。此外,试验中供试的15头取食粉蚧的雌虫都有产卵,而取食幼虫的15头雌虫中有3头在观察的10d内由始至终都没有产卵。
饲料也显著影响孟氏隐唇瓢虫的日均产卵量(图6),取食幼虫的孟氏隐唇瓢虫日均产卵量在试验的前8d内均显著低于取食粉蚧的处理 (t1=-4.147, df=9.231, P(2-tailed)< 0.01;t2=-2.364,df=13,P(2-tailed)=0.034 < 0.05;t3=-3.305,df=12.989, P(2-tailed)< 0.01;t4=-3.962,df=15,P(2-tailed)=0.03 < 0.05;t5=-3.923,df=15,P(2-tailed)=0.016 < 0.05;t6=-3.797,df=16.601, P(2-tailed)< 0.01;t7=-2.854,df=20,P(2-tailed)=0.01 < 0.05;t8=-2.726,df=23,P(2-tailed)=0.012 < 0.05),但是到达第9d、第10 d时两种饲料饲养下的孟氏隐唇瓢虫雌虫日均产卵量差异不显著 (t9=-1.744,df=24,P(2-tailed)=0.094;t10= -1.395, df=22.448,P(2-tailed)=0.177)。
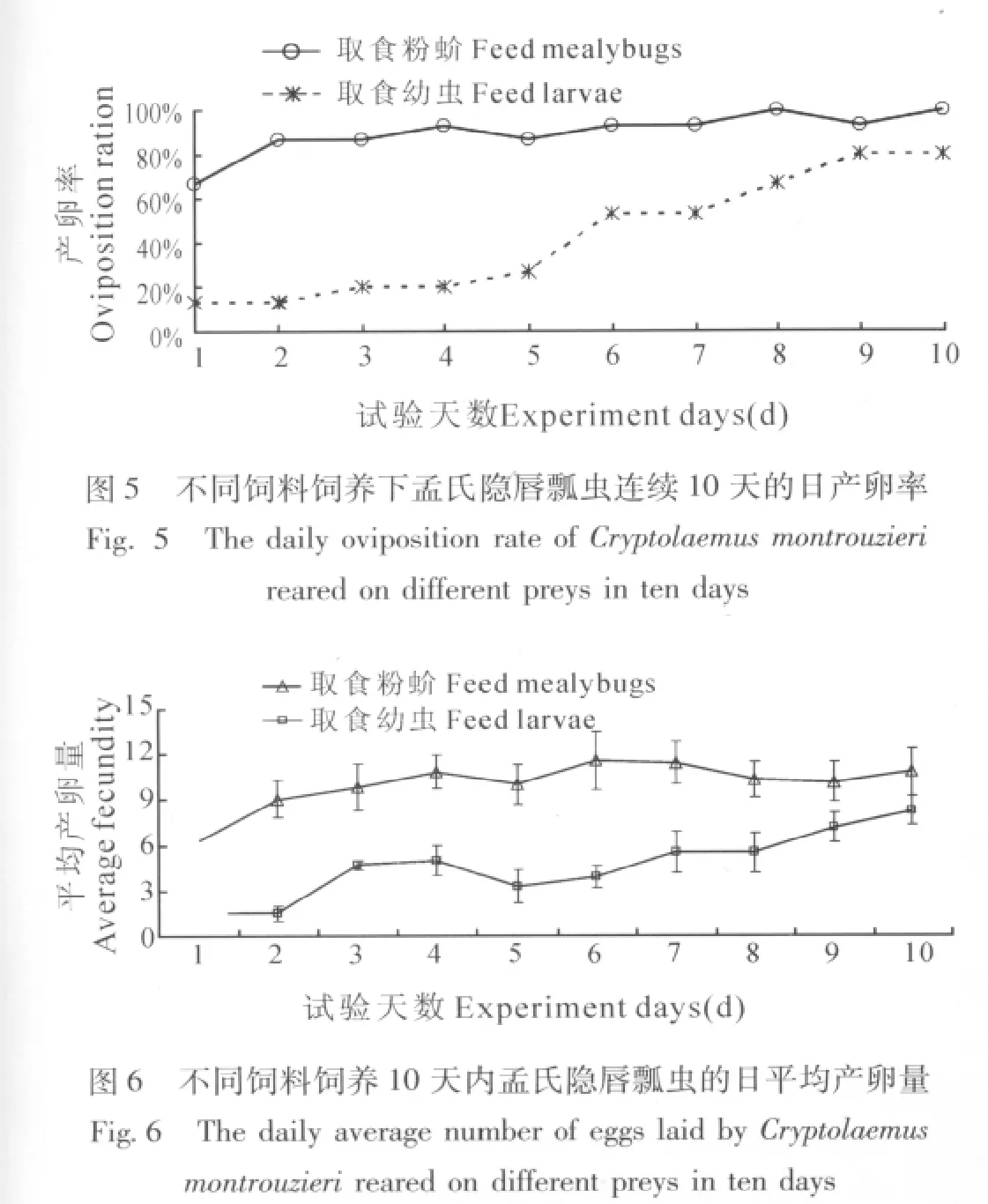
用幼虫饲养孟氏隐唇瓢虫,卵的平均发育历期为5.02 d,以粉蚧饲养的处理为5.09 d,,两个处理无显著差异 (t=-0.383,df=50,P(2-tailed)=0.706),取食这两种饲料的孟氏隐唇瓢虫所产卵的孵化率分别为83.33% (幼虫处理)和90%(粉蚧处理),差异也不显著 (Fisher's exact test:P(2-sided)=0.706)。
3 结论与讨论
我们在试验中发现,孟氏隐唇瓢虫成虫对高龄幼虫的自残强烈程度因成虫性别不同而有差异,雌成虫要明显强于雄成虫。因为雄虫虫体小于雌虫,而瓢虫虫体大小决定着对能量的需求 (Dixon,2007),另外,雌虫强烈的自残习性也可能与其产卵的能量需求相关。从自残过程所持续的时间来看,孟氏隐唇瓢虫雌虫在饱受饥饿时,对低龄幼虫的自残相当厉害,但是幼虫体表形态对自残持续的时间影响显著,正常幼虫由于体表覆盖蜡丝在一定程度上抵御了成虫的攻击,延长了被残杀死亡的时间,然而这种防御在田间有着重要的意义,可以让幼虫在多种捕食者共存的生态位中免于种内自残和种间捕食所造成的死亡,而这种防御意义在小毛瓢虫属Scymnus瓢虫中也曾有报道(Völkl and Vohland,1996)。
雌虫产卵通常会受到同种幼虫足迹的抑制(Merlin et al.,1996;Yasuda et al.,2000;R˚uˇziˇcka,2006),而幼虫足迹中的产卵阻碍信息素 (Oviposition-Deterring Pheromone,ODP)来源于幼虫腹部末端的第十节臀板 (Laubertie et al.,2000),在自残试验中观察到雌虫对低龄幼虫的攻击主要集中在柔软的腹部,除了幼虫腹部体积较大外,这可能还与雌虫能监测到幼虫腹部末端分泌的ODP相关,因为当幼虫挣脱成虫的攻击后,成虫一般会沿着幼虫爬行的轨迹迅速追击。而在异色瓢虫Harmonia axyridis 4龄幼虫的相互残杀中,腹部也是主要的攻击部位 (王甦等,2010)。此外,孟氏隐唇瓢虫幼虫能够利用嗅觉定位猎物 (Sengonca et al.,1995),但对于该瓢虫幼虫能否监测到种内的这种ODP,作为自残时追踪同类的信息还有待进一步研究探讨。
Hodek(1962)把能维持幼虫发育和成虫产卵的食物称之为“基本食物”,仅能作为延长存活的食物称之为“替代食物”。试验中孟氏隐唇瓢虫取食同种幼虫可以产卵,虽然产卵量显著低于取食粉蚧的雌虫,但是这对于成虫渡过逆境,繁衍后代有着重要的适应意义。按照Hodek的理论,同种的幼虫可以作为该瓢虫的基本食物。本试验未探讨取食同种卵对成虫生殖力的影响,但有文献报道龟纹瓢虫Propylea japonica取食自身的卵会丧失繁殖力 (王根和魏建华,1990),这样看来,卵作为食物似乎无法满足瓢虫成虫生殖的需要,只能作为成虫渡过逆境的一种“替代食物”。
瓢虫无论是卵 (Osawa,1989)或是幼虫 (王甦等,2010)的非同血缘自残都比同血缘自残更为严重,因为同血缘相残会减少共有基因转移给下一代,即造成适合度的潜在损失 (Dixon,2000),而种群内个体的行为却是使适合度最大化 (戈峰,2008),所以会尽量避免同血缘自残的发生。本文所涉及的自残均为非同血缘自残,对自残行为的研究可以为该瓢虫的大量扩繁工作提供理论依据和科学指导,虽然孟氏隐唇瓢虫幼虫体表的蜡质能在一定程度上抵御种内自残,但是其成虫在饥饿情况下,尤其是雌虫,会严重威胁到幼虫的存活,因此,成虫大量羽化时需要补充大量的饲料,在其交配产卵后,还应适当转移成虫,避免成虫与幼虫大量共存,造成幼虫死亡。
References)
Agarwala BK,Dixon AFG,1992.Laboratory study of cannibalism and interspecific predation in ladybirds.Ecological Entomology,17(4):303-309.
Dixon AFG,2000.Insect Predator-Prey Dynamics:Ladybird Beetles and Biological Control.Cambridge,United Kingdom:Cambridge University Press.
Dixon AFG,2007.Body size and resource partitioning in ladybirds.Population Ecology,49:45-50.
Fox LR,1975.Cannibalism in natural population.The Annual Review of Ecology,Evolution,and Systematics,6:87-106.
GagnéI,Coderre D,Mauffette Y,2002.Egg cannibalism by Coleomegilla maculatalengi neonates:preference even in the presence of essential prey.Ecological Entomology,27:285-291.
Ge F,2008.Principle and Methods of Insect Ecology.Beijing:Higher Education Press,52.[戈峰,2008.昆虫生态学原理与方法.北京,高等教育出版社,52]
Hodek I,1962.Essential and alternative food in insects.In 11thInternational Congress of Entomology,Vienna(1960),2:696-697.
Laubertie E,Martini X,Cadena C,Treilhou M,Dixon AFG,Hemptinne JL,2006.The immediate source of the oviposition-deterring pheromone produced by larvae of Adalia bipunctata(L.)(Coleoptera,Coccinellidae).Journal of Insect Behavior,19(2):231-240.
Li LY,1993.The research and application prospects of Cryptolaemus montrouzieri in China.Natural Enemies of Insects,15(3):142-152.[李丽英,1993.我国孟氏隐唇瓢虫研究与应用展望.昆虫天敌,15(3):142-152]
Merlin J,Lemaitre O,Gregoire JC,1996.Chemical cues produced by conspecific larvae deter oviposition by the coccidophagous ladybird beetle,Cryptolaemus montrouzieri.Entomologia Experimentalis et Applicata,79:147-151.
Osawa N,1989.Sibling and non-sibling cannibalism by larvae of a lady beetle Harmonia axyridis Pallas(Coleoptera:Coccinellidae)in the field.Researches in Population Ecology,31:153-160.
Pang H,Tang C,Li LY,1996.The technology for preservation of Cryptolaemus montrouzieri adult with the artificial diet.Natural Enemies of Insects,18(2):64-66.[庞虹,汤才,李丽英,1996.孟氏隐唇瓢虫的人工保种技术.昆虫天敌,18(2):64-66]
R˚uˇziˇcka Z,2006.Oviposition-deterring effects of conspecific and heterospecific larval tracks on Cheilomenes sexmaculata(Coleoptera:Coccinellidae).European Journal of Entomology,103:757-763.
Sengonca C,Kotikal YK,Schade M,1995.Olfactory reactions of Cryptolaemus montrouzieri Mulsant(Col.,Coccinellidae)and Chrysoperla carnea(Stephens)(Neur.,Chrysopidae)in relation to period of starvation.Anzeiger Schäedlingskunde Pflanzenschutz Umweltschutz,68:9-12.
Venkatesan T,Singh SP,Jalali SK,2001.Development of Cryptolaemus montrouzieri Mulsant(Coleoptera:Coccinellidae),a predator of mealybugs on freeze-dried artificial diet.Journal of Biological Control,15:139-142.
Völkl W,Vohland K,1996.Wax covers in larvae of two Scymnus species:do they enhance coccinellid larval survival?Oecologia,107:498-503.
Wang G,Wei JH,1990.Studies of behavior of the ladybird beetle Propylea japonica feeding on egg cannibalism.Natural Enemies of Insects,12(3):101-104.[王根,魏建华,1990.龟纹瓢虫残食同种卵行为的研究.昆虫天敌,12(3):101-104]
Wang MQ,Cai WZ,2004.Autotomy in insects.Entomological Knowledge,40(3):280-284.[王孟卿,彩万志,2004.昆虫的肢体自残现象.昆虫知识,40(3):280-284]
Wang S,Tan XL,Zhang F,2010.Influence of kin relationship on cannibalism behavior of Harmonia axyridis(Coleoptera:Coccinellidae)fourth instar larvae in different temperature conditions.Acta Ecologica Sinica,30(19):5396-5403.[王甦,谭晓玲,张帆,2010.不同温度条件下异色瓢虫四龄幼虫亲缘关系对自残行为的影响.生态学报,30(19):5396-5403]
Yasuda H,Takagi T,Kogi K,2000.Effects of conspecific and heterospecific larval tracks on the oviposition behaviour of the predatory ladybird,Harmonia axyridis(Coleoptera:Coccinellidae).European Journal of Entomology,97:551-553.
