磷高效马尾松种源磷效率的家系变异及苗期-大田回溯相关分析
2012-11-21周志春刘伟宏储德裕
杨 青,张 一*,周志春,刘伟宏,储德裕
(1中国林业科学研究院亚热带林业研究所,浙江富阳311400;2浙江省淳安县姥山林场,浙江淳安311700)
磷素是植物生长必需的大量元素,同时又是土壤中有限且不可再生的战略性矿质资源[1-2]。由于土壤对磷素的吸附固定作用,易被植物吸收利用的“活化态”有效磷含量普遍匮乏,已成为全球范围内影响农作物和森林生产力的主要限制性因素。大量研究证实,植物不同基因型之间在磷素吸收和利用效率上存在较大的遗传差异[3-7],因此,充分发掘并利用植物不同基因型对土壤难溶态磷素的生物利用效率的遗传差异,将是提高贫瘠立地条件上植物生产力的有效手段。我国华南地区大面积的丘陵和山地是我国速生人工林的主要分布区和工业用材林的重要基地,然而该地区的酸性红壤有效磷匮乏尤为严重,有效磷含量多在 2 mg/kg[3-4]左右,远不能满足植物正常生长的需求。因而充分挖掘林木吸收利用土壤磷素的遗传潜力、培育磷素营养高效基因型,将成为提高林木对土壤磷素利用效率、实现人工林可持续发展和高产经营的重要途径。
马尾松(Pinus massoniana)人工林面积居我国所有造林树种之冠,是重要的工业用材树种和造林先锋树种。由于马尾松自然分布区内多为有效磷匮乏的酸性红壤,地力衰退和磷素贫瘠已严重制约其人工林生产力。开展马尾松磷效率的遗传改良、提升磷素贫瘠立地上马尾松人工林生产力,将具有重大的育种学和生态学意义[8-15]。目前,相关研究已初步揭示了马尾松磷效率的种源变异规律,并鉴定和筛选出广东信宜、福建武平两个磷效率较高的优良种源[5,10]。同时,已证实马尾松磷效率在家系水平上也存在丰富的遗传变异,在家系水平上开展马尾松磷效率的遗传改良具有较大应用潜力。
然而,当前研究尚存在明显的不足,主要表现为两个方面:一是目前磷效率改良仅分别在种源、家系水平上单独开展[9-12],较少考虑在种源磷效率改良的基础上,深入发掘磷高效种源内部家系水平上的磷效率变异,通过在磷高效种源内部进一步选择磷高效家系,实现“优中选优”、最大限度地发掘马尾松磷效率遗传改良的潜力;第二,对于马尾松等多年生树种而言,通过苗期盆栽试验所鉴定和筛选的磷高效种质,其在大田条件下的多年生长稳定性和磷效率还是未知的,即磷高效基因型早期鉴定的可靠性和稳定性尚未充分证实。因此极为有必要开展苗期磷效率-大田生长回溯相关分析,以验证磷高效基因型苗期选择的可靠性和稳定性,然而,这一研究目前尚未引起重视。鉴于此现状,本试验利用已证实高磷效率的马尾松广东信宜种源为试材,从中随机选择一批代表性的自由授粉家系,并设置不同磷素水平的模拟盆栽试验,旨在获悉磷高效种源内部家系之间磷效率的变异规律,揭示在磷效率种源改良的基础上,进一步开展磷效率家系改良的潜力。在此基础上,结合5年生子代测定林开展马尾松苗期磷效率-大田生长回溯相关分析,揭示开展马尾松磷高效基因型的早期鉴定和选择的可靠性及必要的改良策略。
1 材料和方法
1.1 材料来源
以来自广东信宜种源的9个马尾松优良自由授粉家系作为试验材料,从马尾松一代种子园中随机选择7个优良自由授粉家系的平均性状值,同时以来自马尾松一代种子园的7个自由授粉家系作为对照进行研究,这7个对照家系分别来自不同的典型种源区,具体参见表2和文献[11]。参试家系与对照家系的所有盆栽试验条件均保持一致。盆栽试验所用基质取自千岛湖姥山林场的贫瘠酸性红壤,其有机质含量为6.4 g/kg,全氮和全磷含量分别为0.47 g/kg、0.34 g/kg,水解氮、速效钾和有效磷含量分别为16.2 mg/kg、37.9 mg/kg和1.08 mg/kg,pH 5.07。
1.2 盆栽试验
盆栽试验安排在浙江省淳安县姥山林场试验大棚内进行,于2010年3月播种。基质土壤经风干过筛后,加入1.5% 的钒土(Al2O3)和不同浓度过磷酸钙,利用矾土能吸附并缓慢、均匀释放有效磷的特性,控制介质对有效磷的供应,防止磷从表层介质随水分向下层介质扩散。利用直径12 cm、高度25 cm的塑料筒(定制)作为培养容器。设置低磷(LP)和高磷(H-P)两个养分环境。低磷环境采用上述未加过磷酸钙的基质土壤,高磷环境中过磷酸钙含量为1.0 g/kg。所有处理中,添加适量的氮、磷等元素,维持磷素以外其它元素在正常供应水平,消除其他元素缺乏对马尾松生长的影响。
9个马尾松自由授粉家系按照不同的处理方式采用20次重复的随机区组设计,共计360盆。采用自动喷灌系统维持土壤田间持水量在50%~80%范围内,当土壤水分降低到田间持水量50%左右时喷灌系统开始启动,直至达到田间持水量80%左右为止,这样整个生育期内土壤水分平均为田间持水量的65%左右,以维持马尾松对水分的正常需求。
马尾松家系5年生子代测定林位于浙江省平湖市国营林场,试验为完全随机设计,每个家系20株重复。子代测定林的立地条件为典型的酸性红壤贫磷土壤,其前茬为多年生针阔混交林。本试验所选马尾松样本是在正常生长的植株中随机选取的,舍弃遭受人为或自然因素破坏的植株。最终,每个家系选取12株,依次测定树高、胸径和冠幅等指标,并计算单株材积。
1.3 苗木采收及指标测定
盆栽试验苗于2010年11月采收,不同养分环境中每个家系随机选取15株生长正常的苗木,测量苗高和地径。经清水冲洗后,将苗木分成根、茎、叶3部分。用RHI-ZO Pro STD1600+型根系图像分析系统(加拿大REGENT公司)测定除主根以外的各层次根系长度、根表面积、根系平均直径和根体积等根系形态参数。然后经105℃杀青30min,80℃烘干至恒量,测定茎、叶和根的干物质量,用H2SO4-H2O2消煮—钼锑抗比色法测定各部分的磷含量。
1.4 磷效率的计算和数据分析
相对磷效率、吸收效率和利用效率分别按照如下公式计算:相对磷效率=低磷胁迫下干物质积累量/高磷条件下干物质积累量×100%;磷素吸收效率=磷吸收量/株;磷素利用效率=干物质积累量/磷 吸 收 量[16]。 单 株 材 积 (V)按 公 式V=0.000 062 341 803××估算[13]。
用SAS统计软件的ANOVA程序按家系、磷肥水平分别进行单因素和两因素的方差分析,以检验家系和磷肥效应。采用简单相关分析估算低磷水平下马尾松家系相对磷效率、磷吸收效率和磷利用效率与根构型和生长状况的相关系数,以及与5年生子代测定林的树高、冠幅、胸径及材积的相关性。
2 结果与分析
2.1 马尾松生长和干物质积累量及磷效率的差异
苗高、地径等生长性状和干物质量积累是评价低磷胁迫下马尾松生长适应性和磷效率的重要指标。本试验结果表明,在低磷胁迫条件下,与随机选择的来自不同马尾松典型种源区的一代种子园优良家系(杨青等[11])相比,参试的广东产地马尾松家系总体上表现出较高的干物质生产能力(表1)。9个参试的广东家系的全株干物质积累量平均值为0.75g,比7个对照家系的均值(0.62g)高21%(表2),9个参试家系中有6个家系在低磷胁迫下全株干物质积累量高于对照家系均值。与对照家系相比,参试的9个马尾松家系除5、197和230家系生长性状低于对照外,其它家系都较之有明显的生长优势。进一步分析发现,在低磷胁迫下参试的9个马尾松家系之间在苗高、地茎、叶干重、茎干重、根干重和全株干物质量等主要磷效率性状上的差异都达到了极显著水平(P<0.0001),在参试的9个家系中,以185、337、33和67家系的生长状况最好,其全株干物质积累量平均比9个参试家系的均值高出31.68%,而以5和230家系生长最差。在高磷环境下,除个别性状外,各家系之间的差异均未达到显著水平(表1)。在低磷胁迫下,马尾松不同家系生长性状的遗传变异程度较高磷处理更大,意味着在家系水平上开展马尾松耐低磷胁迫能力和磷效率的遗传改良,具有极大的必要性和改良潜力。
相对磷效率以植物干物质积累量在低磷与正常供磷水平下的比值来表示,这一指标体现了植物对低磷胁迫适应能力的强弱,从侧面则体现了植物对磷肥的敏感程度。从表1来可以看出,9个马尾松家系在低磷胁迫下的磷效率以337和185家系最高(99.48%,95.74%),其次是67、336和33家系,磷效率都达到了86.96%以上。与相对磷效率相反,对磷肥的敏感程度则体现了植物生长对磷肥的反应能力。从磷肥敏感性的角度分析,大部分磷高效家系在低磷胁迫下生长表现优异,但其在低磷、高磷下的生长性状差异不显著,因而其对磷肥的生长反应不敏感(表1)。相反,磷低效的家系如5、197和230则表现出较高的磷肥敏感性,与低磷胁迫相比,其在高磷处理下的生长表现得到显著提升。
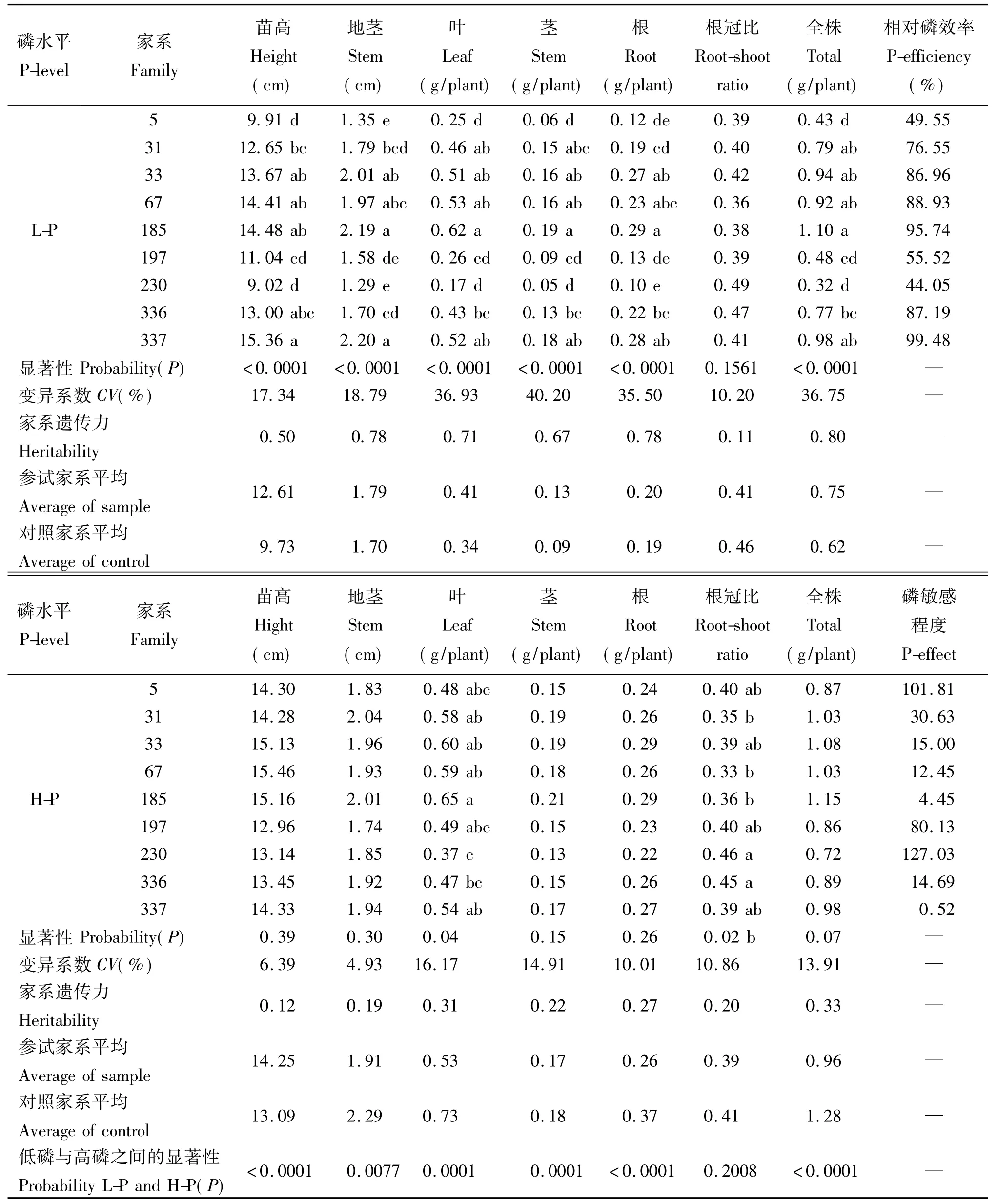
表1 不同磷水平下马尾松9个家系生长性状和干物质积累量Table 1 Differences in growth and dry matter accumulation of 9 families at different phosphorus levels
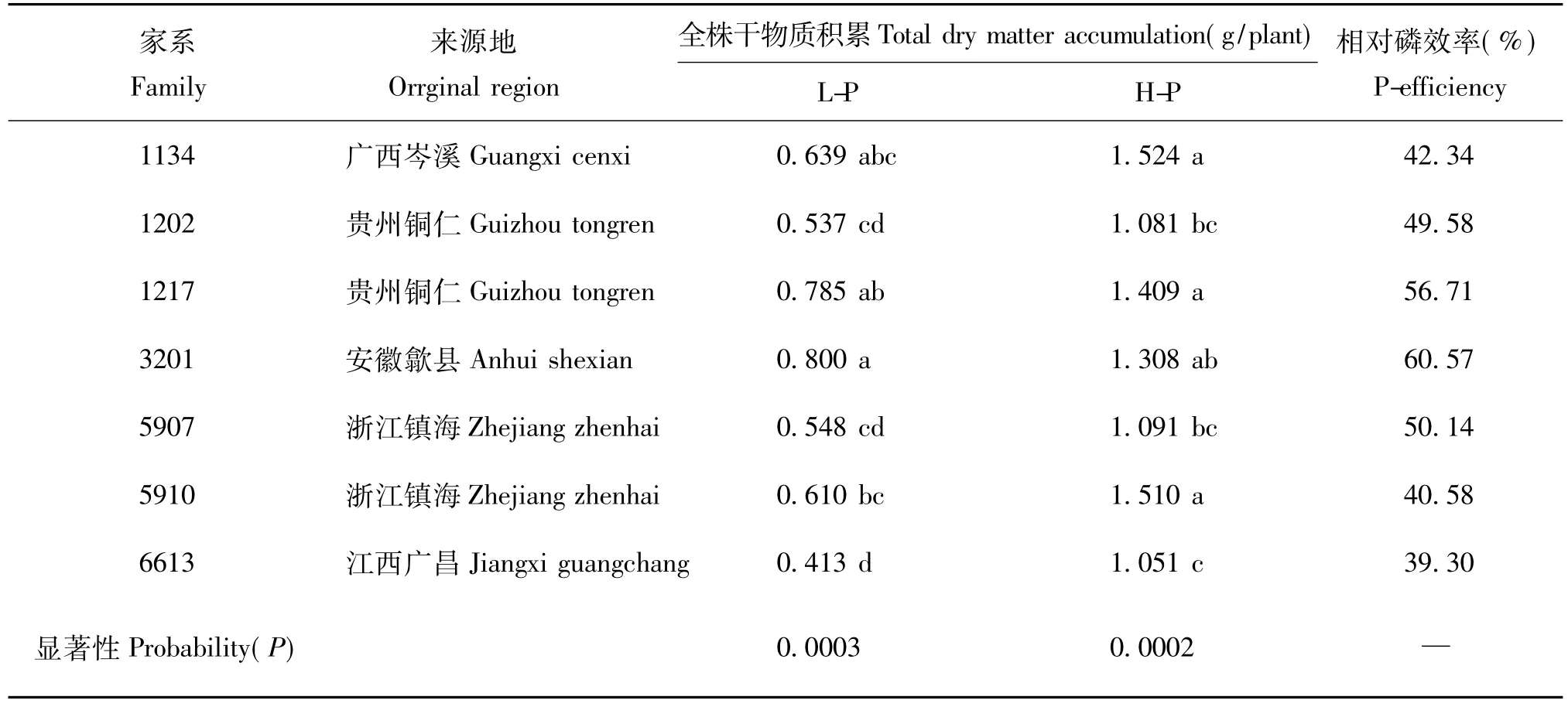
表2 7个对照家系在不同磷水平下的干物质积累量和磷效率Table 2 Dry matter accumulation and phosphorus efficiency of 10 families at different phosphorus levels
2.2 马尾松根系参数分析
表3显示,方差分析显示,在低磷胁迫下马尾松各根系参数在不同家系之间存在着极显著水平的变异(P<0.0001)。在低磷胁迫下,干物质生产能力较强、磷效率较高的家系(185、337、33、67、336和31),其根系长度、根系表面积、根系体积等与其他家系相比均具有较大优势,平均高出其他家系约43.23%(根系长度)、58.55%(根系表面积)和10.87%(根系体积)。家系5和230的根系生长参数较低,而其干物质生产能力和磷效率指标也相对较低。这说明在低磷胁迫下高磷效率马尾松可通过整体根系的增生发育产生较为发达的根系,促进植株对有限养分的吸收和干物质生产能力。在高磷水平下,根系长度、根表面积和根直径也存在显著的家系差异(P<0.0341),其中以33、185、336和337家系根型参数指标最为优良,这与低磷胁迫下的试验结果基本一致。可见这几个家系不仅具有较强的耐低磷胁迫能力,在高磷下也能有效地吸收土壤中的营养成分,继而在干物质生产能力上得到有效的提高。
2.3 磷吸收效率和磷利用效率的家系变异
从表4可以看出,在低磷胁迫下磷效率较高和干物质量生产能力较强的337、185、67和33家系,其整株的磷吸收效率也较高(≥1.36mg/株),其中337家系最高,达到了1.45 mg/株。方差分析结果显示,在低磷胁迫下,马尾松各器官以及全株的磷吸收效率都存在极显著的家系差异(P<0.0001),而在高磷环境下家系变异较小,各器官及整株的差异均未达到显著水平。
磷利用效率以单位磷素的植株干物质积累量表示,反映的是植物对所吸收磷素在植株体内的运转、再分配和利用能力。方差分析结果显示,在低磷胁迫下马尾松叶、根及整株的磷素利用效率的家系变异达到了显著性水平(P=0.0195)。总体上,磷效率高的家系具有较高的磷素吸收效率,而低磷效率家系其磷素吸收效率也较低,与磷高效家系的差异达到极显著水平(表4),例如磷效率最高的4个家系33、67、185和337,其磷素吸收效率均超过了1.3 mg/株,而磷效率较低的家系5、197和230,其磷素吸收效率均低于0.60 mg/株。进一步分析发现,磷效率高的家系不仅具有较高的磷素吸收效率,其磷素利用效率也较高,而低磷效率家系的磷素利用效率则相应较低。例如,磷效率最高的家系33、67、185、337其磷素利用效率分别达到了1.41、1.47和1.23和1.46 g/mg,而磷效率最低的5和230家系其磷素利用效率分别为1.16和1.17 g/mg。然而,与磷低效家系相比,磷高效家系在磷素吸收效率上的优势要明显大于其在磷素利用效率上的优势。
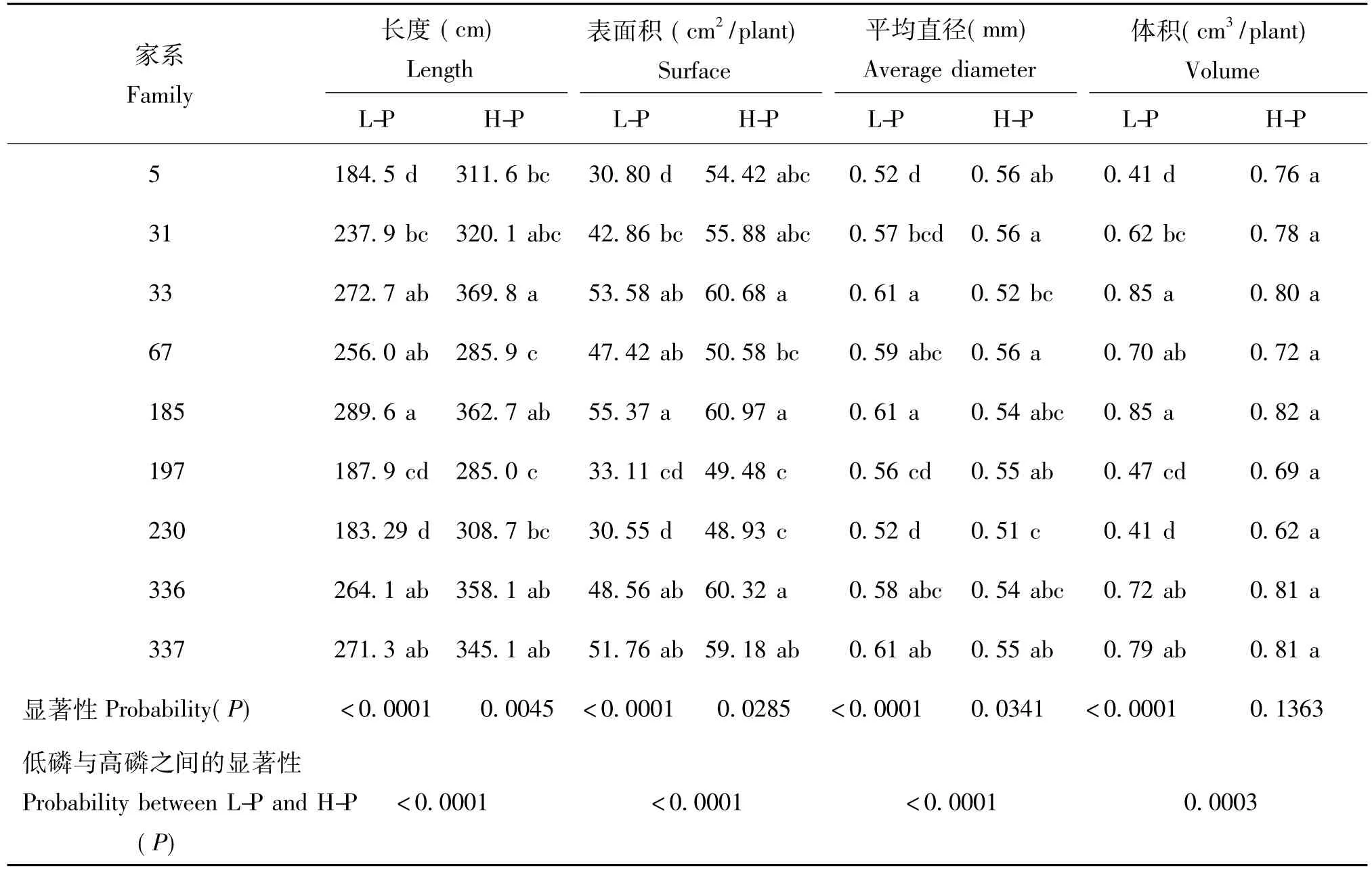
表3 不同磷水平下马尾松各家系根系参数Table 3 Difference of root morphological parameter at different phosphorus levels
2.4 马尾松家系磷效率子性状相关分析
相关分析(表5)发现,马尾松家系所有的磷效率子性状,包括低磷胁迫下的植株苗高、地径、干物质积累量,与磷素吸收效率的相关性均达到了极显著水平(r=0.9783)。相比较,磷效率子性状中仅苗高与磷素利用效率的相关性达到了显著水平,而其它磷效率子性状与磷素利用效率的相关系数虽大于0.6,却未达到显著水平。同时,马尾松各磷效率子性状与主要根系形态参数,包括根系长度、根表面积、根系体积等均呈极显著水平的正相关,且相关系数均大于0.90。
2.5 马尾松家系盆栽磷效率性状与5年生家系测定林生长性状相关分析
利用与盆栽试验相同材料的5年生马尾松家系子代测定林,进行了苗期-大田生长回溯相关分析,以验证通过开展磷效率苗期盆栽试验鉴定选育高磷效率马尾松家系的可行性。从总体上看(表6),参试马尾松家系在苗期低磷胁迫条件下的生长性状,即苗期磷效率指标,与低磷立地大田中5年生测定林的生长性状在一定程度上一致。在低磷胁迫盆栽试验中表现出较高或中等以上磷效率的家系,如185、336、67、33,其在大田条件下也生长较优。而在苗期盆栽试验中磷效率较低的家系5、230,其在大田条件下生长性状也较差。不过,也有部分家系的苗期磷效率性状与大田生长表现不一致,如家系197在盆栽试验中磷效率较低,而在大田试验中其材积性状值较高;家系337的苗期磷效率较高,而在大田试验中材积值较低,仅为0.0049。相关分析发现(图1),马尾松家系在苗期低磷胁迫下的生长性状即磷效率指标,与其在大田条件下树高、胸径和材积等性状一般呈正相关,但不同性状的相关程度不一,其中苗期的主要磷效率指标与大田条件下的树高性状均呈显著或极显著水平(P<0.05)相关,而苗期主要磷效率指标,即植株生物量与大田条件下材积性状仅达到0.10水平的相关(P<0.1)。
以上结果表明,马尾松家系的苗期生长状况和磷效率指标能在一定程度上预测大田生长条件下马尾松家系的生长适应能力和生产力,然而这种预测的准确程度还有待于进一步研究和验证。
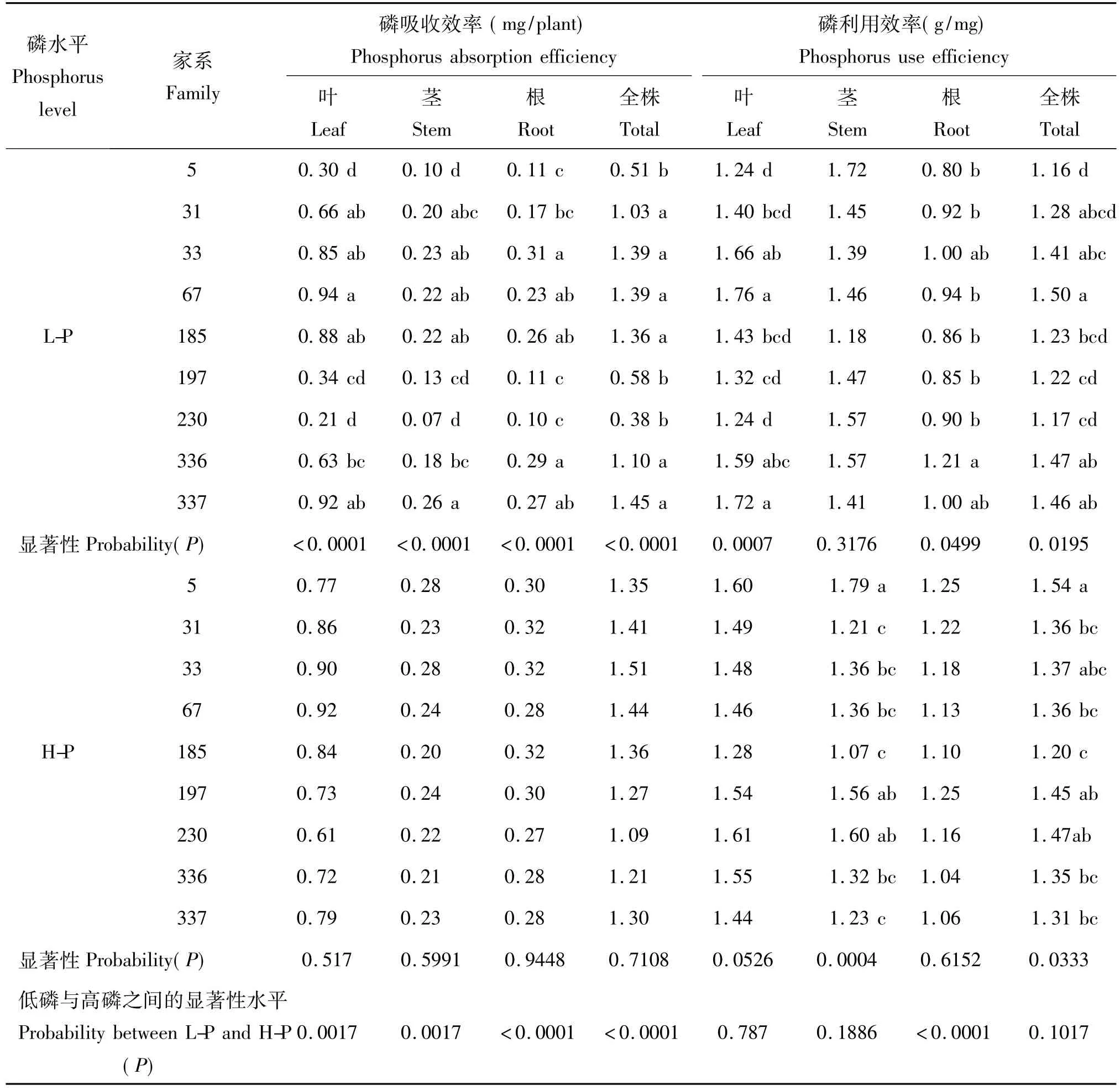
表4 不同养分环境下马尾松家系的磷吸收效率和利用效率Table 4 Phosphorus absorption efficiency and phosphorus use efficiency of 9 families at different phosphorus levels
3 讨论
已有的研究揭示,马尾松是磷效率在种源和家系水平上均存在丰富的遗传变异,证实磷效率具有较大的遗传改良潜力[8-14]。然而,目前马尾松磷效率改良仅分别在种源、家系水平上单独开展[9-12],而在磷效率的种源改良基础上,在磷高效种源内部深入发掘家系水平上的磷效率变异,以及在磷高效种源内选择磷高效家系,并通过“优中选优”最大限度地发掘马尾松磷效率的遗传潜力等方面还缺少研究。本文研究结果表明,来自广东信宜种源的自由授粉家系在低磷胁迫下的干物质生产力和磷效率,总体上高于作为对照的马尾松一代种子园优良家系,进一步证实了该种源具有较高的磷效率和生长适应性。同时,分析发现参试家系的苗高、地茎、干物质积累量等主要磷效率指标在家系间的变异均达到显著或极显著水平,揭示了在马尾松高磷效率优良种源内部进一步发掘并充分利用磷效率的家系变异,选育磷高效优良家系具有较大的潜力和可行性。分析表明,主要磷效率指标的家系遗传力均大于0.70,进一步证实了选育高磷效率马尾松家系的改良潜力。
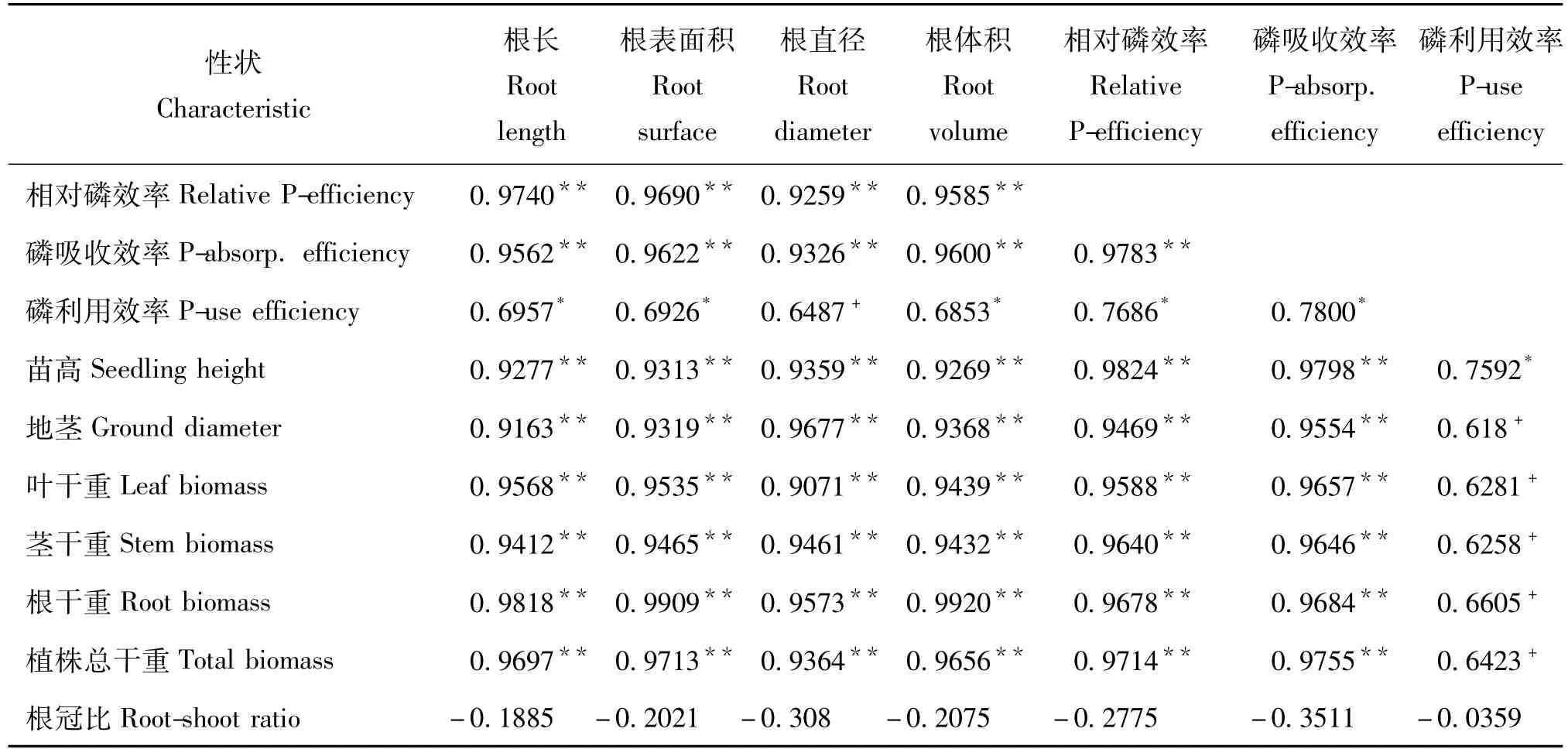
表5 低磷胁迫下马尾松磷效率子性状之间的表型相关系数Table 5 Correlation between phosphorus efficiency traits of 9 families
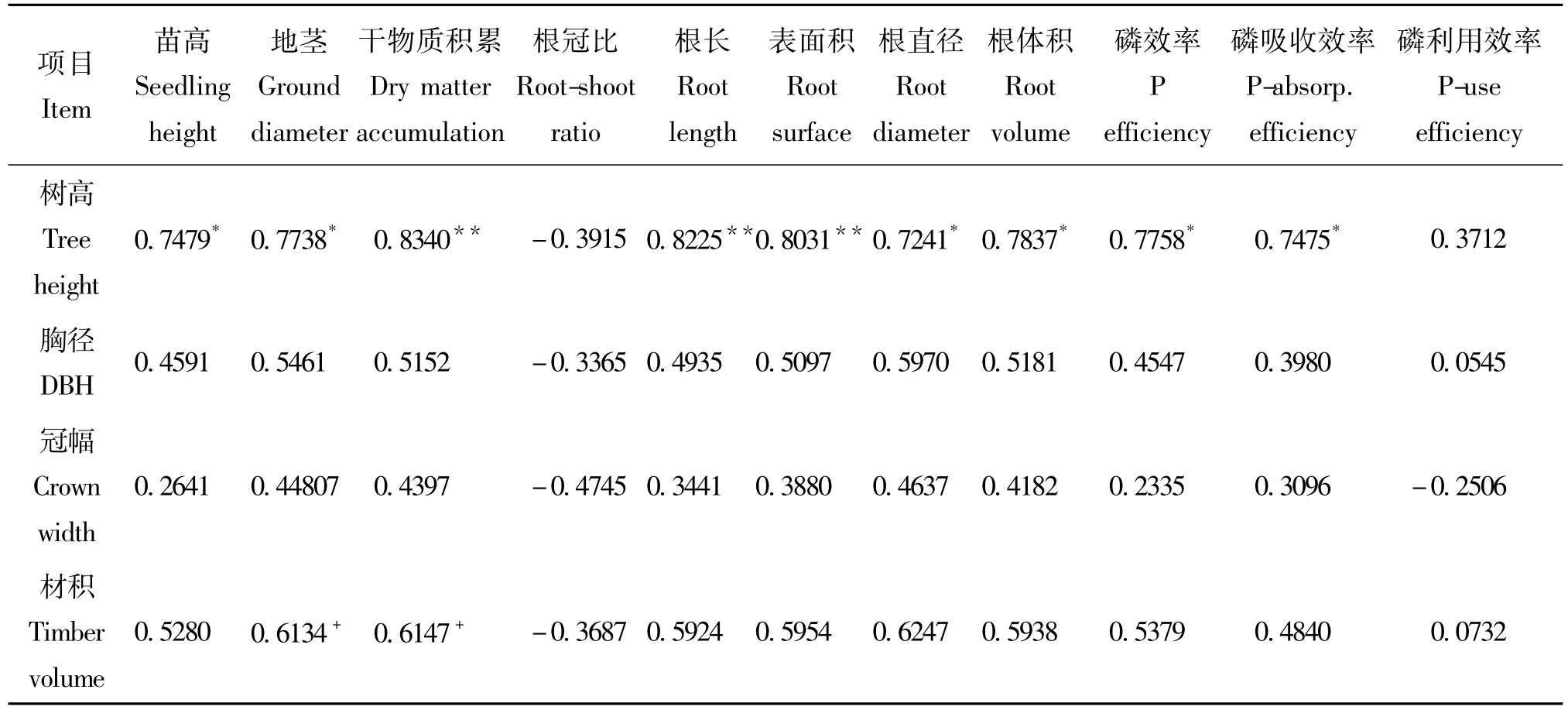
表6 苗期生长性状及磷效率与大田生长性状回溯相关分析Table 6 Correlation between growth traits in seedling and 5-year-age progeny
已有研究揭示,低磷胁迫下根系的适应性生长发育和磷素吸收效率的增加,以及植物对所吸收磷素利用效率的提高是作物不同品种适应低磷胁迫、高效获取磷素的重要形态和生理基础[17-21]。磷吸收效率和磷利用效率对于植物磷效率的相对重要性,一般因不同植物种类或基因型而异[10,22-23],磷素吸收效率高的植物,一般通过根系形态和生理适应性变化增加对土壤中磷素的获取,而磷素利用效率较高的植物,则更多通过增加磷素在植物体内的高效转运和合理利用提高其磷效率[1,24-26]。本文研究揭示,磷高效马尾松家系在低磷胁迫下的根系长度、根系总表面积等参数均显著高于低磷效率家系。同时,发现马尾松磷效率与磷素吸收效率表现显著的正相关,而磷效率与磷素利用效率则相关程度较低。这一结果揭示,高磷效马尾松家系主要通过根系形态参数的适应性变化,产生了较大的根系长度、根系体积和根系表面积,增加了根系与土壤的接触面积、增强了对土壤中大量难溶态磷的活化、吸收和利用,最终实现了较高的磷效率和生产力。
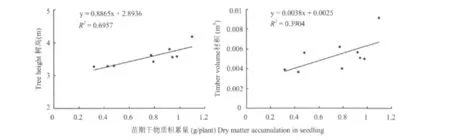
图1 苗期生物量与大田测定林树高、材积线性相关图Fig.1 Correlogram of biomass,tree height and timber volume of seedlings of progeny tested forestry
对于马尾松等多年生树种而言,通过苗期盆栽试验所鉴定和筛选的磷高效种质,其在大田条件下的后期生长稳定性和磷效率还是未知的,因此开展苗期磷效率-大田生长回溯相关分析极为必要[27]。本文通过苗期-大田生长回溯相关分析发现,最重要的苗期磷效率指标—干物质积累量与大田条件下的树高性状呈显著相关(P<0.05),表明通过马尾松家系的苗期磷效率指标,能较好地预测大田生长条件下马尾松的树高等生长性状。不过,苗期生物量与材积性状仅达到0.10水平的相关(P<0.1),表明苗期盆栽试验-大田生长回溯相关的程度尚有进一步提高的必要。
在挪威云杉和欧洲赤松中研究发现,苗期-成熟期生长相关性较低,其主要原因是人工智能温室与大田试验的环境因子不一致[16,28-29]。本文中马尾松苗期低磷胁迫盆栽试验在开放式的条件下开展,其水、热条件均与大田试验条件同步,同时,本论文中盆栽试验所用基质为有效磷极低的酸性红壤,与开展大田试验的土壤肥力接近。据推断,这是本实验中苗期-大田回溯相关的程度高于挪威云杉和欧洲赤松中相关研究结果的主要原因。森林土壤中磷素养分的空间分布具有多样性和复杂性[11],本研究中大田试验立地的前茬为多年生针阔混交林,由于来自阔叶树的凋落物分解较为迅速,可能会导致土壤表层有效磷浓度较高、而深层土壤有效磷浓度较低,从而形成较为复杂的异质低磷胁迫。而本文中低磷胁迫模拟的盆栽试验,则均以土壤磷素呈均一分布为前提开展,导致盆栽磷素养分环境与森林土壤中的异质低磷环境不完全一致。据推断,这应该是导致本研究中部分性状的苗期-大田回溯相关性不高的主要原因。针对这一问题,今后可对大田试验森林土壤中有效磷的浓度和空间分布特征进行全面检测,在此基础上,完全按照自然条件下的磷素养分空间分布特征开展盆栽实验,这样将有望提高苗期-大田生长性状回溯相关的程度,更高效地开展马尾松高磷效率种质的早期鉴定和选育。
4 结论
1)磷高效种源内不同家系的磷效率指标在家系间的变异显著,在马尾松高磷效率优良种源内部进一步发掘并充分利用磷效率的家系变异,选育磷高效优良家系具有较大的潜力和可行性。
2)苗期磷效率指标与大田试验中树高性状呈显著相关(P<0.05)相关,通过马尾松家系的苗期磷效率指标能较好地预测大田生长条件下马尾松的树高等生长性状。
3)苗期磷效率指标与大田试验中材积性状相关不显著,应检测自然森林土壤中的磷素养分空间分布特征并据此开展盆栽实验,以提高苗期-大田生长性状回溯相关的程度。
[1]Vance C P.Symbiotic nitrogen fixation and phosphorus acquisition.Plant nutrition in a wet tropical forest[J].Ecosystem,2004,7:404-419.
[2]Beebe S E,Pierce M R,Yan X L.Quantitative trait loci for root architecture traits correlated with phosphorus acquisition in common bean[J].Crop Sci.,2006,46:413 -423.
[3]李庆奎.中国红壤[M].北京:科学出版社,1985.145-146.Li Q K.Chinese soil[M].Beijing:Science Press,1985.145-156.
[4]邱燕,张鼎华.南方酸性土壤磷素化学研究进展[J].福建稻麦科技,2003,21(3):14-17.Qiu Y,Zhang D H.Research progress on phosphorus transformation in southern acid soils[J].Fujian Sci.Technol.Rice Wheat,2003,21(3):14-17.
[5]周志春,谢钰容,金国庆.马尾松种源对磷肥的遗传反应及根际土壤营养差异[J].林业科学,2003,39(6):62-67.Zhou Z C,Xie Y R,Jin G Q.Genetic response of Pinus massoniana provenances to phosphorus supply and nutrient characteristics of their rhizosphere soil[J].Sci.Silv.Sin.,2003,39(6):62 -67.
[6]严小龙.热带土壤中菜豆种质耐低磷特性的评价[J].植物营养与肥料学报,1995,1(1):30-37.Yan X L.Evaluation of bean germplasm for low phosphorus tolerance on tropical soils[J].Plant Nutr.Fert.Sci.,1995,1(1):30-37.
[7]沈宏,施卫明,王校常,曹志洪.不同作物对低磷胁迫的适应机理研究[J].植物营养与肥料学报,2001,7(2):172-177.Shen H,Shi W M,Wang X C,Cao Z H.Study on adaptation mechanisms of different crops to low phosphorus stress[J].Plant Nutr.Fert.Sci.,2001,7(2):172 -177.
[8]谢钰容,周志春,金国庆,等.低P胁迫对马尾松不同种源根系形态和干物质分配的影响[J].林业科学研究,2004,17(3):272-278.Xie Y R,Zhou Z C,Jin G Q et al.Root morphology and dry matter allocation of masson pine:Response of different provenances to low phosphorus stress[J].For.Res.,2004,17(3):272 -278.
[9]谢钰容,周志春,廖国华,等.低磷胁迫下马尾松种源酸性磷酸酶活性差异[J].林业科学,2005,41(3):58-62.Xie Y R,Zhou Z C,Liao G H et al.Difference of induced acid phosphate activity under low phosphorus stress of Pinus massoniana provenances[J].Sci.Sil.Sin.,2005,41(3):58 -62.
[10]周志春,谢钰容,金国庆,等.马尾松种源磷效率研究[J].林业科学,2005,41(4):25-30.Zhou Z C,Xie Y R,Jin G Q et al.Study on phosphorus efficiency of different provenances of Pinus massoniana[J].Sci.Sil.Sin.,2005,41(4):25-30.
[11]杨青,张一,周志春,等.异质低磷胁迫下马尾松家系根构型和磷效率的遗传研究[J].植物生态学报,2011,35(12):1226-1235.Yang Q,Zhang Y,Zhou Z C et al.Genetic voriation in root architecture and phosphorus efficiency in response to heterogeneous phosphorus deficiency in pinus massoniana families[J].Chin.J.Plant Ecol.,2011,35(12):1226 -1235.
[12]周志春,谢钰容,金国庆,等.马尾松磷效率及相关性状的家系遗传和变异[J].北京林业大学学报,2004,26(6):1-5.Zhou Z C,Xie Y R,Jin G Q et al.Inheritance and variation of phosphorus efficiency and its related traits in families of Pinus massoniana[J].J.Beijing For.Univ.,2004,26(6):1 -5.
[13]周志春,傅玉狮,吴天林.马尾松生长和材性的地理遗传变异及最优种源区的划定[J].林业科学研究,1993,6(5):556-564.Zhou Z C,Fu Y S,Wu T L.Geographic variation in growth and wood propeties and detemination of theoptimum provenance region for paper-pulp wood of masson pine[J].For.Res.,1993,6(5):556-564.
[14]赵颖,周志春,吴吉富,等.马尾松优良种源对磷肥的生长反应和肥效持续性[J].林业科学,2007,43(10):64-70.Zhao Y,Zhou Z C,Wu J F et al.Growth response of superior provenance of mason pine to phosphorus supply and persistence of phosphorus effect[J].Sci.Sil.Sin.,2007,43(10):64 -70.
[15]梁霞,刘爱琴,马祥庆,等.磷胁迫对不同杉木无性系酸性磷酸酶活性的影响[J].植物生态学报,2005,29(1):54-59.Liang X,Liu A Q,Ma X Q et al.The effect of phosphorus deficiency stress on activities of acid phosphatase in different clones of Chinese fir[J].J.Plant Ecol.,2005,29(1):54 -59.
[16]曹靖,张福锁.低磷条件下不同基因型小麦幼苗对磷的吸收和利用效率及水分的影响[J].植物生态学报,2000,24(6):731-735.Cao J,Zhang F S.Phosphorus uptake and utilization efficiency in seedlings of different wheat genotypes as influenced by water supply at low soil phosphorus availability[J].Acta Phytoecol.Sin.,2000,24(6):731-735.
[17]孙海国,张福锁.缺磷胁迫下的小麦根系形态特征研究[J].应用生态学报,2002,13(3):295-299.Sun H G,Zhang F S.Morphology of wheat roots under lowphosphorus stress[J].Chin.J.Appl.Ecol.,2002,13(3):295-299.
[18]孙海国,张福锁,杨军芳.不同供磷水平小麦苗期根系特征与其相对产量的关系[J].华北农学报,2001,16(3):98-104.Sun H G,Zhang F S,Yang J F.Characteristics of root system of wheat seedlings and their relative grain yield[J].Acta Agric.Bor.-Sin.,2001,16(3):98 -104.
[19]赵华,徐芳森,石磊,等.植物根系形态对低磷胁迫应答的研究进展[J].植物学通报,2006,23(4):409-417.Zhao H,Xu F S,Shi L et al.Advances in plant root morphology adaptability to phosphorus deficiency stress[J].Chin.Bull.Bot.,2006,23(4):409 -717.
[20]任海红,刘学义,李贵全.大豆耐低磷胁迫研究进展[J].分子植物育种,2008,6(2):316-322.Liu H H,Liu X Y,Li G Q.Advances of soybean tolerant to low phosphorus stress[J].Molec.Plant Breed.,2008,6(2):316 -322.
[21]何文寿,陈素生,康建宏.宁夏春小麦氮素利用效率的基因型差异研究[J].土壤,2003,35(6):500-505.He W S,Chen S S,Kang J H.Genotypic differences in nitrogen recovery rate between spring wheat varieties in Ningxia[J].Soils,2003,35(6):500-505.
[22]梁霞,刘爱琴,马祥庆,等.不同杉木无性系磷素特性的比较[J].植物生态学报,2006,30(6):1005-1011.Liang X,Liu A Q,Ma X Q et al.Comparison of the phosphorus characteristics of different Chinese fir clones[J].J.Plant Ecol.,2006,30(6):1005-1011.
[23]Pierret A,Moran C J,Doussan C.Conventional detection methodology is limiting our ability to understand the roles and functions of fine roots[J].New Phytol,2005,166:967 -980.
[24]梁泉,廖红,严小龙.植物根构型的定量分析[J].植物学通报,2007,24(6):695-702.Liang Q,Liao H,Yan X L.Quantitative analysis of plant root architecture[J].Chin.Bull.Bot.,2007,24(6):695 -702.
[25]刘灵,廖红,王秀荣,等.不同根构型大豆对低磷的适应性变化及其与磷效率的关系[J].中国农业科学,2008,41(4):1089-1099.Liu L,Liao H,Wang X R et al.Adaptive changes of soybean genotypes with different root architectures to low phophorus avaiability as related to phosphorus efficiency[J].Sci.Agric.Sin.,2008,41(4):1089-1099.
[26]Zhang Y,Ma X H,Zhou Z C.Foraging ability and growth performance of four subtropical tree species in response to heterogeneous nutrient environments[J].J.For.Res.,2010,15(2):91-98.
[27]Jansson G,Jonsson A,Eriksson G.Use of trait combinations for evaluating juvenile - mature relationships in Picea abies(L.)[J].Tree Gen.& Genom.,2005,1:21-29.
[28]Sonesson J,Jansson G,Eriksson G.Retrospective genetic tests of Pinus sylvestris L.in growth chambers with two ir-rigation regimes at two temperatures[J].Scand J.For Res,2001,6:21-29.
[29]Sonesson J,Jansson G,Eriksson G.Retrospective genetic testing of Picea abies under controlled temperature and mois-ture regimes[J].Can J.For.Res,2002,32:81 -91.
