水稻单穗质量和千粒质量的遗传分析
2012-11-10陈小荣陈志彬贺浩华朱昌兰彭小松贺晓鹏傅军如欧阳林娟
陈小荣,陈志彬,贺浩华,朱昌兰,彭小松,贺晓鹏,傅军如,欧阳林娟
(江西农业大学农学院,作物生理生态与遗传育种教育部重点实验室,江西省作物生理生态与遗传育种重点实验室,江西南昌 330045)
单穗质量作为水稻Oryza sativa L.穗部性状的重要指标,越来越受到重视.张声函等[1]研究指出,穗质量是株高、有效穗数、穗实粒数、结实率、百粒质量、穗质量等主要经济性状指标中影响单株生产力的最主要因素.事实上,一般认为选育大穗型品种是水稻超高产或超级稻育种的主要手段[2-4].前人对大穗型品种的基本特征进行了大量研究[5-8],遗传上研究也较多[9-14].粒质量是构成水稻产量的重要因子,水稻品种的粒质量不但对其产量有直接影响,而且对品质也有间接影响[15~17].籽粒大小多以千粒质量计[18].董桂春等[19]研究表明千粒质量是影响穗质量的重要因子之一,特别对于一些大穗型品种,其千粒质量均稳定在较高的水平.前人对千粒质量的遗传也进行了一些研究[9-14,18,20-26],但由于研究材料与分析方法(遗传模型)的不同,所得结论存在差异.
由于数量性状遗传研究分析上的复杂性,稻作界对水稻产量构成因素遗传研究需进一步深入.近年来,盖钧镒等[27-28]提出了1套主基因+多基因混合遗传分析方法并建立了相应的遗传模型,得到较好的应用[29-30].本研究选择单穗质量和千粒质量差异大的3个亲本配制2个杂交组合,建立相应的P1、F1、P2、B1、B2和 F26 个世代,并将 2 套遗传群体分为中、晚2个生产季节种植,利用该套遗传分析方法对单穗质量和千粒质量进行了遗传分析,目的是探讨水稻穗粒质量的遗传特性,以期为水稻穗粒遗传改良提供参考.
1 材料与方法
1.1 材料
本研究采用的水稻材料CB1、CB4以及CB7均是江西农业大学作物遗传育种研究室选育的农艺性状稳定的常规稻,其单穗质量分别在1.21、3.46和1.42 g左右,千粒质量分别在22.2、32.3和19.4g左右,3个亲本材料中单穗质量和千粒质量均以CB4最大,CB1与CB7均远低于CB4,彼此差异显著.
选择上述亲本材料,配制CB1×CB4和CB7×CB4 组合,通过自交和杂交等建立 P1、F1、P2、B1、B2和F26个世代,将种子随机分为2份:1份中季种植,1份晚季种植.
1.2 方法
1.2.1 性状调查 材料种植于江西农业大学科技园试验田.中季于2009年5月20日播种,6月20日移栽;晚季于6月22日播种,7月22日移栽.株行距18 cm×18 cm,单本种植,常规栽培管理.亲本取40株,F1代取30株,回交世代分别取120株,F2代取200株,按照常规法分别测单穗质量、有效穗数和千粒质量等.
1.2.2 数据分析方法 应用盖钧镒[27]等提出的植物数量性状主基因+多基因混合遗传模型分析方法,对 P1、F1、P2、B1、B2和 F2等 6 个世代进行遗传分析.通过极大似然法和IECM(Iterated expectation and conditional maximization)算法对混合分布中的有关成分分布参数做出估计,然后通过AIC值的判别和一组适应性检验,选择最优遗传模型,并估计主基因和多基因效应值、方差等遗传参数.
2 结果与分析
2.1 单穗质量
2.1.1 亲本和组合群体在各世代中的单穗质量表1表明,单穗质量较小的亲本与较大亲本杂交组合F1单穗质量接近甚至大于大值亲本,表现出极强的杂种优势.B1、B2回交世代平均值可看出,与大值亲本回交,其后代平均穗质量相对于小值亲本回交后代要高,如CB7/CB4组合群体晚季种植下B2平均穗质量是B1的1.399倍.2个群体2个季别下F2世代的穗质量平均值均介于2个亲本之间,趋于中间值.

表1 单穗质量在各世代中平均数Tab.1 Mean of grain mass per panicle in all generations
2.1.2 模型的选择及遗传参数的估算 单穗质量各模型的AIC值见表2,选取AIC值最小及与最小AIC值最接近的2个遗传模型,如CB1/CB4组合群体中季种植下,AIC值最小的为D-2模型,与之最接近的是C-0模型和D-1模型,选择该3个模型进行适合性检验,包括:均匀性检验()、Smirnov检验(nW2)、Kolmogorov检验(Dn),选择统计量达到显著水平个数较少的模型作为最优模型.
表3列出了各遗传群体穗质量的各种遗传模型的AIC值.由表3可知,D-2模型在检验中达到显著水平的个数较其他2个模型的少,因此可以确定D-2模型为CB1/CB4组合群体中季种植下最适模型.2个组合群体通过各种模型的检验,选出最适合模型,并求出模型参数极大似然估计值,结果列于表4.经过检验,2个组合群体2个种植季节下单穗质量最佳模型均为D模型(一对基因+多基因模型).通过最小二乘法估算,求出一阶遗传参数估计值,列于表5.由表5可知,各群体单穗质量的群体均值(m)分别为2.4584、2.5786、2.7543 和2.8481,主基因的加性效应值(d)分别为 -0.4640、-0.7066、-0.3901和 -0.3359.
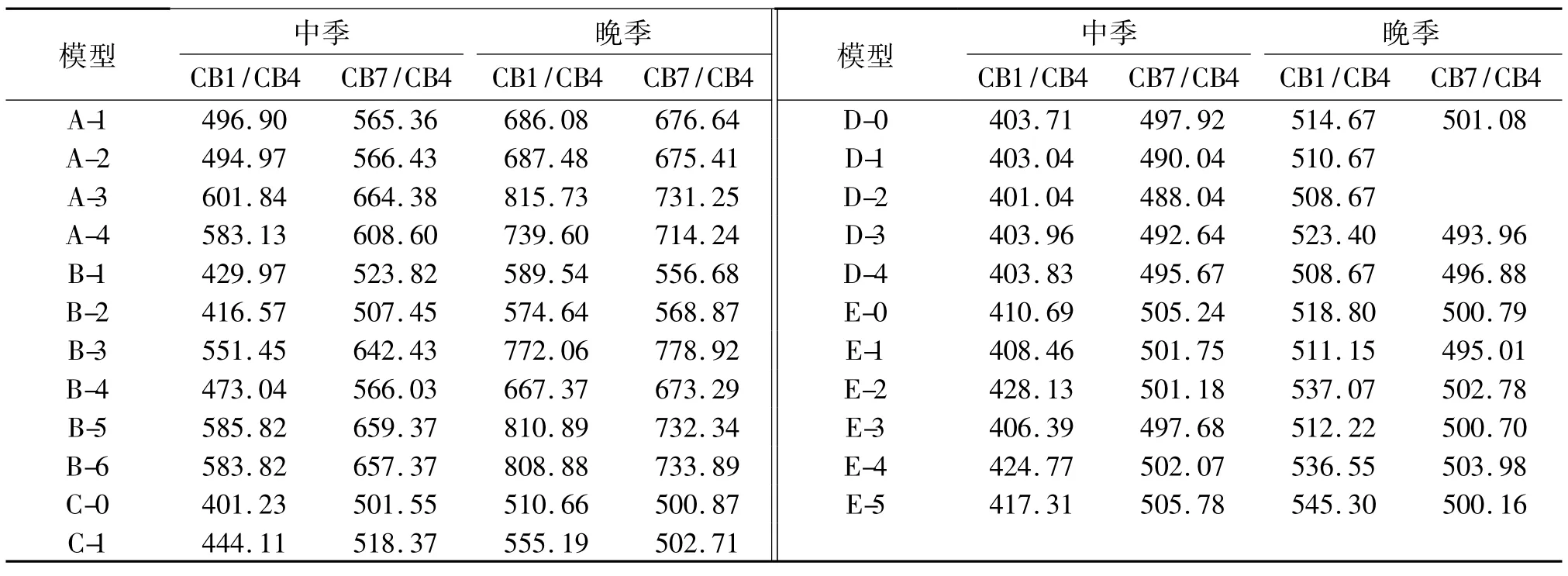
表2 各群体单穗质量各种遗传模型的AIC值Tab.2 The AIC values of grain mass per panicle in the all populations under various genetic models
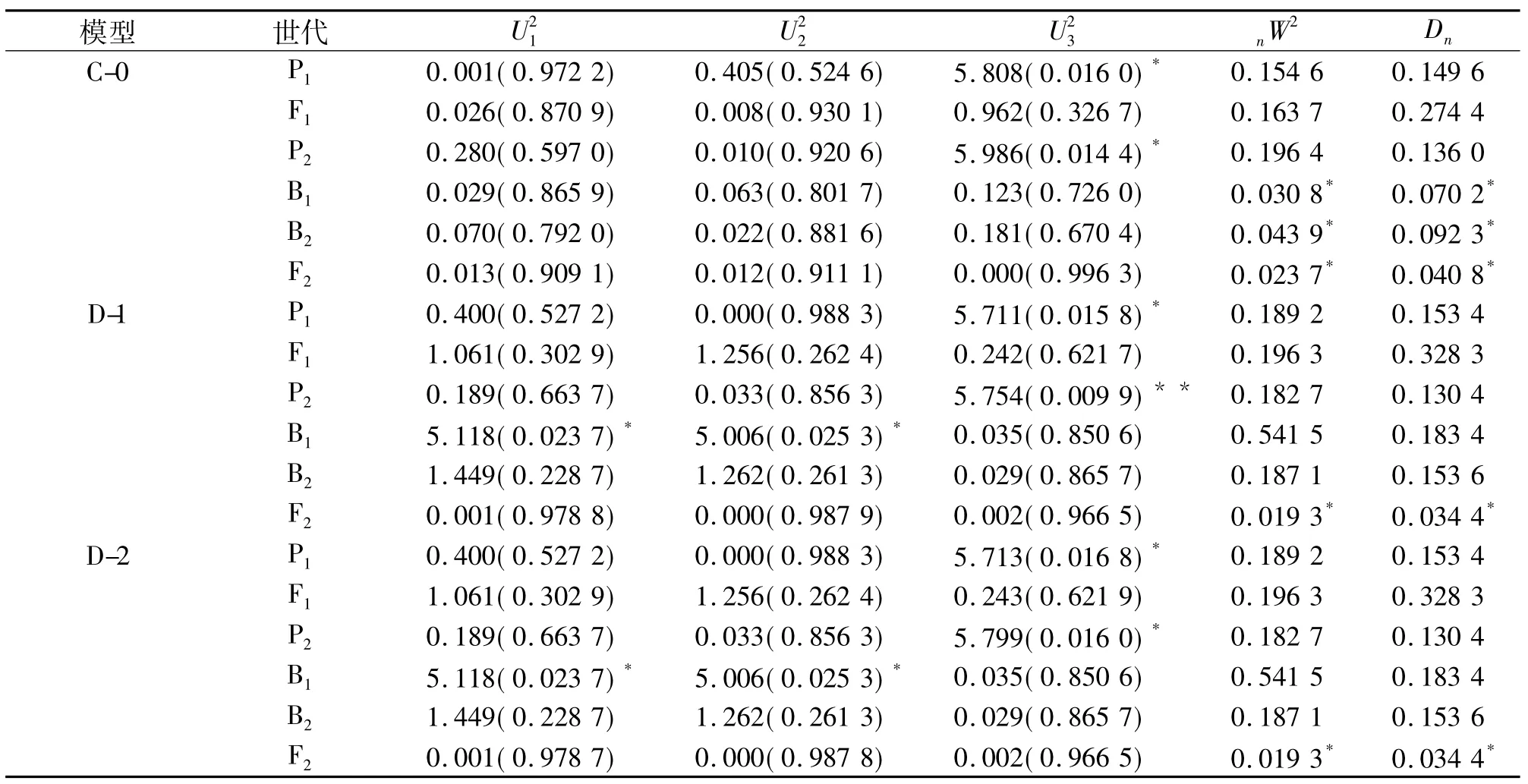
表3 CB1/CB4组合群体中季种植下部分模型的适合性检验1)Tab.3 Test for goodness of fitness of population of combination CB1/CB4 as middle season planting models

表4 各个世代最佳模型及模型参数极大似然估计值1)Tab.4 The best model and the maximum likelihood estimates in the all generations
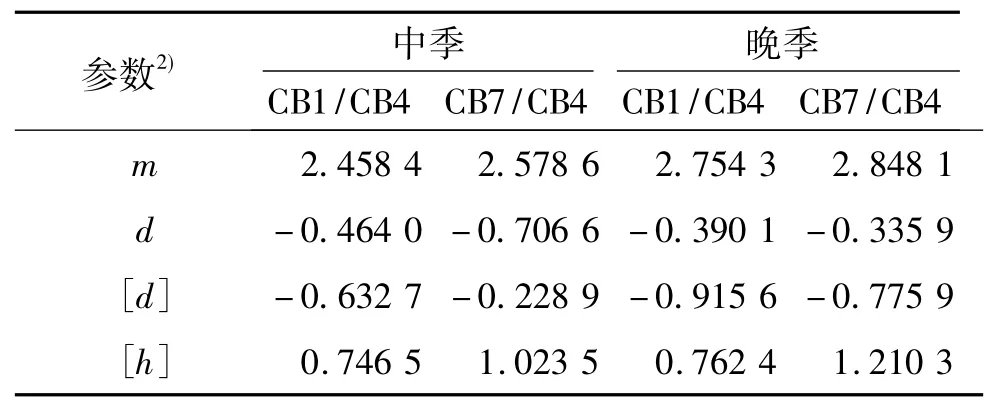
表5 各群体单穗质量一阶遗传参数的估计值1)Tab.5 The first order parameter estimates of the grain mass per panicle of the all populations
进一步求出二阶遗传估计值,列于表6.由表6可以看出,CB1/CB4组合群体中季种植下主基因遗传率为58.06%~69.23%,多基因遗传率为18.36%~25.46%,总基因型遗传率为 79.51% ~87.60%;CB7/CB4组合群体中季种植下主基因遗传率为58.27%~62.68%,多基因遗传率为5.39% ~23.60%,总基因型遗传率为68.07% ~81.88%;CB1/CB4组合群体晚季种植下主基因遗传率为69.81%~75.24%,多基因遗传率为5.03% ~21.13%,总基因型遗传率为80.27% ~92.60%;CB7/CB4组合群体晚季种植下主基因遗传率为59.36%~75.60%,多基因遗传率为20.86% ~23.74%,总基因型遗传率为83.10% ~96.68%.
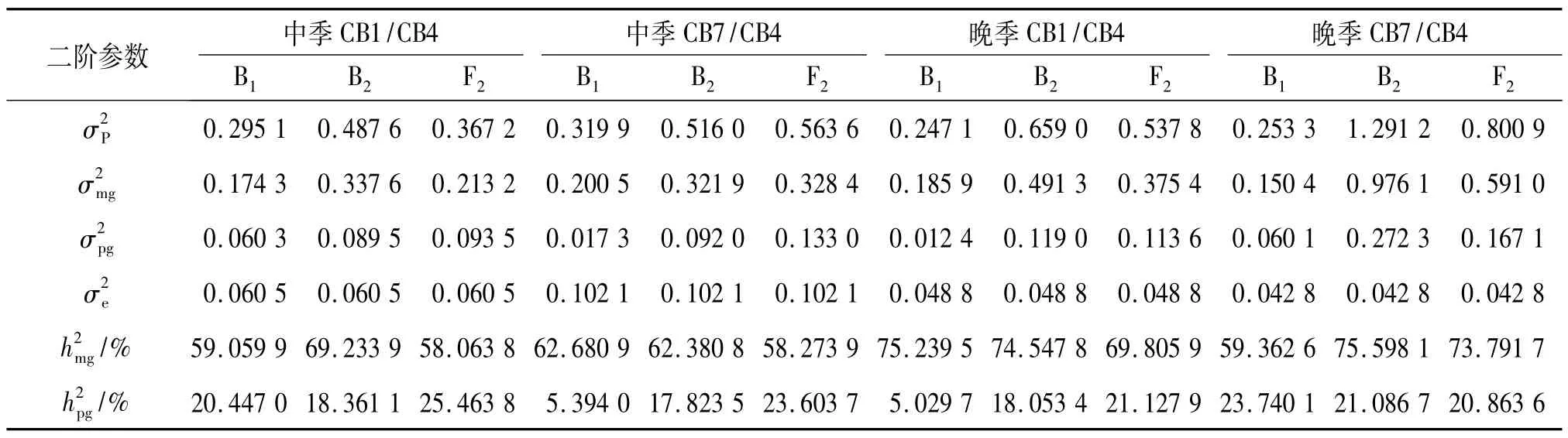
表6 各群体单穗质量的二阶遗传参数1)Tab.6 The second order parameter estimates of the grain mass per panicle of the all populations
2.2 千粒质量
与单穗质量模型选择程序相似,经过检验,2个群体2个种植季节下千粒质量最佳模型均为D-2模型(加性主基因+加-显性多基因模型).通过最小二乘法估算,求出一阶遗传参数估计值,列于表7,由表7可知,各群体千粒质量群体均值(m)分别为29.2468、27.2353、29.1221 和 27.5447,主基因加性效应值(d)分别为 -2.6186、-1.2524、-1.1711和 -1.1977.
进一步求出二阶遗传估计值(表8),表8表明,CB1/CB4组合群体中季种植下主基因遗传率为60.06% ~69.39%,多基因遗传率为10.73% ~23.21%,总基因遗传率为71.48% ~83.55%;CB7/CB4组合群体中季种植下主基因遗传率为19.04% ~36.17%,多基因遗传率为51.67% ~70.82%,总基因遗传率为78.71%~92.68%;CB1/CB4组合群体晚季种植下主基因遗传率为28.46% ~30.23%,多基因遗传率为43.08%~52.82%,总基因遗传率为72.57% ~81.28%;CB7/CB4组合群体晚季种植下主基因遗传率为17.59%~34.49%,多基因遗传率为30.38% ~68.89%,总基因型遗传率为64.88% ~86.48%.
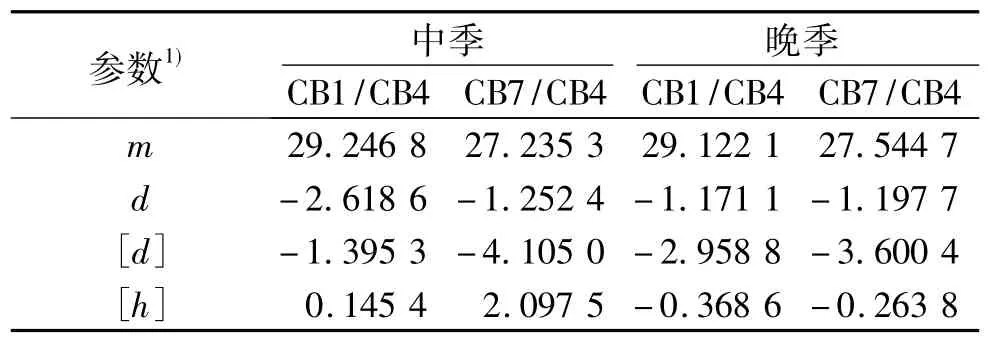
表7 各群体千粒质量一阶遗传参数的估计值Tab.7 The first order parameter estimates of the 1000-grain mass of the all populations
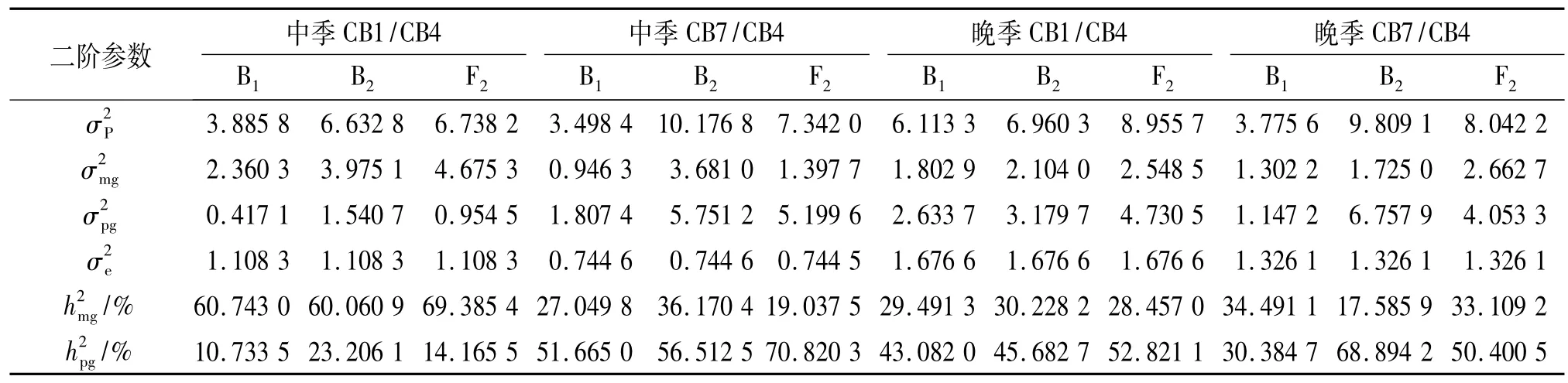
表8 各群体千粒质量的二阶遗传参数1)Tab.8 The second order parameter estimates of the 1000-grain mass of the all populations
3 讨论与结论
单穗质量是水稻最重要的农艺性状之一,杨春华等[13]研究指出单穗质量广义遗传力较高和狭义遗传力中等,以加性效应或加性效应与非加性效应共同作用为主.吴家胜等[14]研究认为单穗质量普通遗传率和互作遗传率分别为25.5%和24.5%,单穗质量QTL主效应和QTL与环境互作效应的联合贡献率分别为37.7% 和29.3%.本试验对3个单穗质量相差较大的材料进行6个世代联合分析得出该性状属于一对主基因控制+多基因模型.B1、B2回交世代对比可发现,大值亲本回交世代单穗质量平均值高于小值亲本回交后代,2个季节均表现出该特征,表明单穗质量的遗传基础可能与细胞质效应有关.同一遗传群体不同季别下对比可发现种植季别对单穗质量遗传率影响较小;但不同组合群体同一季别下存在差异,即CB7/CB4组合群体基因型遗传率中、晚两个季节均比CB1/CB4组合群体大.因此在对单穗质量性状进行遗传研究时,应考虑用于构建群体的亲本因素.进一步对一阶遗传参数分析可发现,主基因加性效应对穗质量的表达有降低作用,选育重穗型品种时应注意之.
千粒质量直接影响水稻品种的丰产潜力,尤其水稻品种产量达到一定水平后,千粒质量将为新品种进一步高产担负越来越重要的角色,因此千粒质量开始受到遗传学家和育种学家重视.陈光辉等[18]研究指出,千粒质量性状遗传以基因的加性效应为主.程本义等[16]发现千粒质量在年度间、不同环境条件下相对稳定遗传.王松凤等[21]则认为粒形相关性状与千粒质量的QTL受主效基因控制的同时,也受环境影响.石春海等[22]研究认为谷粒的长度、宽度、厚度、长度比和粒质量均属于多基因控制的数量性状,有较高的遗传力.蔡英杰[23]研究表明千粒质量受1对主基因+多基因控制,主基因决定千粒质量41.29%表型变异,多基因决定48.60%.欧海龙[24]研究则表明千粒质量属于2对隐性上位主基因+多基因控制遗传模型,主基因遗传率为79.0%,多基因遗传率为12.4%.江建华等[25]研究也表明千粒质量受2对主基因+多基因控制,且2对主基因是连锁的.吴家胜等[14]检测到15个QTI控制千粒质量性状,其中12个QTL有加性效应,6对QTL有加加上位性效应,环境条件对其无影响.本试验对3个千粒质量各异的材料分中、晚季2个温光环境种植,进行6个世代联合分析,均得出千粒质量属于一对加性主基因+加-显性多基因模型的D-2模型,与蔡英杰[23]研究结果较为相似.进一步对B1、B2回交世代对比可发现,大值亲本回交世代千粒质量平均值高于小值亲本回交后代,2个季节均表现出该特征,表明千粒质量的遗传基础可能与细胞质效应有关.本研究千粒质量主基因的加性效应值(d)均为负值,说明主基因的加性效应会降低千粒质量性状的表达,以CB1/CB4组合群体晚季种植下最为明显.CB1/CB4组合群体中季种植时主基因遗传率最大,多基因遗传率则相对较小,一阶参数也是CB1/CB4组合群体中季种植时d为最小.其余主基因、多基因遗传率均较接近,d大小也相近.说明,千粒质量遗传研究时需要考虑用于构建遗传群体的亲本及种植季别的选择.
[1]张声函,雷雪方,朱普年.水稻穗重对高产育种的效应研究[J].江西农业大学学报,1995,17(4):386-389.
[2]周开达,马玉清,刘太清,等.杂交水稻亚种间重穗型组合选育[J].四川农业大学学报,1995,13(4):403-407.
[3]杨守仁,张步龙,陈温福,等.超高产水稻育种的理论和方法[J].中国水稻科学,1996,10(2):115-120.
[4]袁隆平.杂交水稻超高产育种[J].杂交水稻,1997,12(6):1-3.
[5]严进明,翟虎渠,张荣铣,等.重穗型杂种稻光合和光合产物运转特性研究[J].作物学报,2001,27(2):261-266.
[6]马均,周开达,马文波,等.重穗型杂交稻穗颈节间维管束与籽粒充实关系的研究[J].中国农业科学,2002,35(5):576-579.
[7]马文波,马均,明东风,等.不同穗重型水稻品种剑叶光合特性的研究[J].作物学报,2003,29(2):236-240.
[8]马均,马文波,明东风,等.重穗型水稻株型特性研究[J].中国农业科学,2006,39(4):679-685.
[9]MEI H W,XU J L,LI Z K,et al.QTLs influencing panicle size detected in two reciprocal introgressive line(IL)populations in rice(Oryza sativa L.) [J].Theor Appl Genet,2006,112:648-656.
[10]XING Y Z,TAN Y F,HUA J P,et al.Characterization of the main effects,epistatic effects and their environmental interactions of QTLs on the genetic basis of yield traits in rice[J].Theor Appl Genet,2002,105:248-257.
[11]LI Z K,PINSON S R M,PARK W D,et al.Epistasis for three grain yield components in rice(Oryza sativa L.)[J].Genetics,1997,145:453-465.
[12]ZHUANG J Y,FAN Y Y,RAO Z M,et al.Analysis on additive effects and additive-by additive epistatic effects of QTLs for yield traits in a recombinant inbred line population of rice[J].Theoretical and Applied Genetics,2002,105:1137-1145.
[13]杨春华,李仕贵.籼型三系杂交水稻不育系主要农艺性状配合力及遗传力分析[J].西南农业学报,2005,18(3):234-239.
[14]吴家胜,周鸿凯,陈国波,等.水稻单穗重和千粒重的QTL定位[J].中国水稻科学,2008,22(2):143-147.
[15]熊振民,蔡洪法.中国水稻[M].北京:中国农业科技出版社,1992.
[16]程本义,杨仕华,沈伟峰,等.中国长江流域新育成籼稻品种分析及育种应用[J].中国农学通报,2004,20(2):92-96.
[17]李贤勇,何永歆,李顺武,等.水稻千粒重与垩白粒率的相关性分析[J].西南农业学报,2003,16(4):20-23.
[18]陈光辉,周清明,王建龙,等.两系杂交水稻千粒重的遗传研究[J].热带作物学报,2007,28(4):57-61.
[19]董桂春,王余龙,吴华.水稻品种间穗重构成因子的差异性研究[J].扬州大学学报:农业与生命科学版,2003,24(4):62-66.
[20]程本义,沈伟峰,杨仕华.中国南方水稻品种的粒重及其与主要性状间的关系[J].杂交水稻,2007,22(2):73-77.
[21]王松凤,李辉,刘喜,等.水稻粒形相关性状及千粒重QTL的稳定性分析[J].南京农业大学学报,2008,31(3):1-7.
[22]石春海,申宗坦.早籼粒形的遗传和改良[J].中国水稻科学,1995,5(1):27-32.
[23]蔡英杰.水稻长穗大粒RIL群体产量相关性状的遗传分析[D].福州:福建农林大学,2006.
[24]欧海龙.水稻产量和穗部性状主基因+多基因混合遗传[D].福州:福建农林大学,2005.
[25]江建华,洪德林,郭媛,等.粳稻穗角与谷粒性状的相关性及谷粒性状遗传分析[J].植物学报,2009,44(2):167-177.
[26]周清元,安华,张毅.水稻子粒形态性状遗传研究[J].西南农业大学学报,2000,22(2):102-104.
[27]盖钧镒,章元明,王建康.植物数量性状遗传体系[M].北京:科学出版社,2003.
[28]王建康,盖钧镒.利用杂种世代鉴定数量性状主基因-多基因混合遗传模型并估计其遗传效应[J].遗传学报,1997,24(5):432-440.
[29]杨空松,陈小荣,傅军如,等.东乡野生稻育性恢复性的鉴定与遗传分析[J].中国水稻科学,2007,21(5):487-492.
[30]尹建华,刘宜柏,曾红权,等.水稻光温敏核不育系的育性遗传分析[J].生物数学学报,2007,22(1):107-112.
