脑内高表达microRNAs的时空效应及其在神经管缺陷中的可能作用*
2012-11-06白宝玲李会莉陈淑媛王晓蕾赵慧智
白宝玲, 李会莉, 陈淑媛, 王晓蕾, 张 勤, 赵慧智, 张 霆
(首都儿科研究所, 北京 100020)
1000-4718(2012)12-2291-06
2012-07-16
2012-09-20
国家自然科学基金资助项目(No.31071133) ;北京市科委科技新星资助项目(No.2010B045)
△通讯作者 Tel: 010-85695585; E-mail: zhangtingcv@yahoo.com.cn
·综述·
脑内高表达microRNAs的时空效应及其在神经管缺陷中的可能作用*
白宝玲, 李会莉, 陈淑媛, 王晓蕾, 张 勤, 赵慧智, 张 霆△
(首都儿科研究所, 北京 100020)
微小RNA(microRNA,miRNA)在生物体的发育生长、细胞增殖分化及凋亡、激素的分泌和肿瘤发生等领域中具有重要的调控功能[1-2]。哺乳动物中枢神经系统的发育过程需要经历多个位点的神经管闭合,若神经管闭合失败或早闭,会导致神经管缺陷(neural tube defects, NTD)的发生,这是一种严重的、多因素的先天性复杂疾病,以往文献报道有多种miRNAs参与了NTD的发病机制。
1 神经管的形成以及NTD发生的机制
1.1神经管的发育形成 哺乳动物中,神经管闭合被认为是个不连续的过程,该闭合过程沿头-尾轴类似于拉拉链的方式进行分段闭合,如小鼠胚胎中,闭合启动大约在妊娠日(gestational day,GD)8.5 d,从闭合点1、2、3依次逐渐闭合[3-4]。但在人类胚胎中,缺乏闭合点2的形成,且约有80%小鼠胚胎在缺乏该位点后颅骨仍闭合完全[3,5],因此推断此闭合位点对脑的形成是非必需的。
神经管形成由细胞迁移、增殖、黏附、凋亡以及细胞融合和重塑等系列事件组成。研究证明,多条信号通路参与其中,如平面细胞极性(planar cell polarity,PCP)通路所驱动的汇聚延伸对闭合位点1的形成及闭合意义重大,Shh(Sonic hedgehog)通路则调控着神经褶中线铰链位点(median hinge point,MHP)及背外侧铰链位点(dorsolateral hinge point,DLHP)的形成,Notch通路被认为参与了神经干细胞的自我修复和抑制神经元早熟等事件,另外,经典Wnt通路、肌醇代谢通路、维甲酸(retinoic acid, RA)代谢通路、信号转导子和转录激活子通路(signal transducer and activator of transcription,STAT)及骨成型蛋白通路(bone morphogenetic protein, BMP)等都参与了神经系统的形成,且这些通路之间成网状互相协同作用,精密地共调控了神经管的完美闭合。
1.2NTDNTD是由神经管闭合不全或早闭所导致的一种严重先天畸形,临床表现为无脑儿、脊柱裂、露脑畸形、脑膨出、脑脊髓膜膨出等。研究表明NTD的发生是环境和遗传因素共同作用的结果。其中,叶酸代谢途径是研究最为广泛的与NTD发病相关的功能机制,机体缺乏叶酸时可通过减少核酸合成(嘌呤和胸苷),使同型半胱氨酸水平升高及改变体内甲基化水平来引起NTD[6]。此外,在神经管闭合过程中,神经褶形态稳定所需的细胞骨架蛋白、神经褶闭合所需的整合/黏附蛋白、神经细胞不断增殖所需的细胞周期、细胞存活或凋亡相关蛋白、细胞外基质蛋白等这些蛋白因子的功能受到干扰时可能会诱发NTD[5]。而一些转录协同激活因子、染色质重塑复合物、转录调控分子、蛋白质功能调控因子、泛素化降解复合物等,以及前述PCP、Shh等一些重要信号通路的异常,亦都可能诱导NTD的发生。而作为转录后调控基因表达的重要方式之一,miRNAs在神经管形成的各个重要环节中发挥着关键作用,本综述重点对这种调控机制及其对NTD形成的可能机制进行探讨。
2 miRNAs与神经管发育
miRNAs在神经系统发育过程中具有表达的组织特异性或富集性及时空特异性,研究发现,在人和小鼠中,miR-9、-124a、-124b、-135、-153、-183及-219只在大脑组织有特异性的表达,而miR-9*、-125a、 -125b、-128、-132、-137及 -139则在大脑中富集性表达[7]。利用MiRXplore芯片筛选发现,在小鼠胚胎的神经管发育过程中约有72个miRNAs有表达,进一步对处于神经管闭合期GD-8.5、GD-9.0和GD-9.5的胚胎进行miRNAs表达分析发现,有16个miRNAs在三期都有表达,而miR-638只特异在GD-9.0表达,miR-335、-130a、-199a-3p、-18b、-18a及miR-106a特异在GD-9.5表达,GD-8.5则无特异性的miRNA表达[8]。这些miRNAs时空特异性表达的非一致性,可能与大脑等中枢神经系统不同脑区形成中的时空特性具有相对应的特异性调控功能。
在脑内特异性表达的miRNAs中,miR-124及miR-9是研究最为广泛的。成熟的miR-124序列从蠕虫到人类完全保守,在分化或成熟的神经元细胞中特异性高表达[9],它在神经管发育时可能通过抑制神经祖细胞的某些基因表达来维持神经元的分化[10]。如多聚嘧啶核苷酸序列结合蛋白(polypyrimidine tract binding protein, PTBP)1是负调控PTBP2的一种选择性剪接抑制剂,在非神经元细胞或神经祖细胞中高表达。miR-124可调控PTBP1使其表达水平下降,从而使PTBP2的蛋白表达迅速积累并促使非神经细胞向神经细胞的转化,有趣的是,PTBP2也能被miR-124抑制,但抑制效率小于其对PTBP1的抑制效率[11],这或许能解释为什么在神经细胞中PTBP2比PTBP1的表达要迟缓一些。miR-124对神经发育调控的另一个证据来源于对羧基端结构域磷酸酶(small C-terminal domain phosphatases 1,SCPl)的研究,已知SCP1是RE1-沉默转录抑制子(repressor element-1-silencing transcription factor, REST)复合物的组成之一,在非神经元细胞中阻遏神经元特异基因的转录,而miR-124可靶作用SCP1从而解除其对神经元转录程序的阻遏,但同时,miR-124又受到REST的抑制,形成一个双重负反馈环路[12-13]。此外,miR-124还可作用于整合信号蛋白Itgβ1、Itgα7、Itgα3、Itgα11等,影响神经管闭合时神经上皮细胞的迁移和黏附[14-15]。Leucht等[16]发现,中后脑边界处(midbrain-hindbrain boundary,MHB)可释放成纤维生长因子(fibroblast growth factor,Fgf)来诱导位于其前部的神经管发育为前脑,后部的发育为后脑,且转录因子Her5和Her9是维持该区未分化状态的关键分子,而这些分子皆可被 miR-9特异性调控,但miR-9却组织特异性地在MHB以外周围区域表达,使MHB区域内的靶基因免受miR-9的抑制性调控,从而帮助划定了MHB的边界。值得注意的是,miR-124和miR-9能共调控参与许多生物过程的发生,如当神经发育进行时,二者可靶向抑制染色质重塑复合物BAF(Brahma-related gene 1-associated factors)53a的表达,从而使BAF53b得以激活并诱导树突的形成,但在非神经元细胞中,REST沉默了miR-124和miR-9的表达, BAF53a成功抑制BAF53b令细胞维持在神经祖细胞状态[17]。不仅如此,miR-124和miR-9还可共靶作用于STAT3信号通路分子,通过影响肝细胞生长因子(hepatocyte growth factor,HGF)或c-Met,对胚胎干细胞向神经细胞的定向分化进行调控[18-19]。
miR-103与miR-107虽在多种组织和器官中都有表达,但在大脑中表达最高[20]。研究发现,miR-103与miR-107可作用于CDK5R1基因的3’-UTR,使其不能编码出激活CDK5所需的活化剂p35,从而影响神经细胞的迁移[21]。锌指同源蛋白家族(zinc finger protein homologous family,ZH)X1也是接受miR103/107调控的一个潜在靶基因,ZHX1在体外体内实验中均可有效沉默DNA甲基转移酶(DNA methyltransferase, DNMT)3B[22],也就是说,miR103/107间接参与了DNA甲基化的表观修饰,与其协同发挥对神经管发育的调节作用。
已知血管内皮生长因子A(vascular endothelial growth factor A,VEGFA)通路调控高等脊椎动物胚胎的血管生成,而miR-126可负调控神经管发育中VEGFA mRNA的表达,从而对神经系统血管网络的形成和模式化起间接的调控作用[23]。通过拮抗Notch信号通路来实现神经管细胞迁移和血管形成的表皮生长因子(epidermal growth factor,EGF)L7也是miR-126的潜在靶基因[24-25],另外,胰岛素受体底物1(insulin receptor substrate 1,IRS-1)是类胰岛素生长因子(insulin-like growth factor,IGF)通路上重要的成分和SHH通路上一重要的效应分子[26],可通过在神经营养蛋白通路中发挥其对早期胚胎形成和神经管发生中不可或缺的功能作用[27],而miR-126对IRS-1和可诱导IGF-1受体表达的Hoxa9基因都有靶调控作用[28-30],即miR-126可能协同调节着神经管发育过程中IGF、Shh和神经营养蛋白通路的相互作用。
其它miRNAs在神经发育中也起到重要的调控作用,如miR-199a-5p 及miR-199a-3p可靶作用于Brm基因(在发育中的神经管中检测有表达),从而间接影响Swi-Snf染色质重塑复合物对转录激活和抑制的调控作用[31],有趣的是,该2个miRNAs的表达又受到涉及颅神经管闭合的Twist-1蛋白的调节[32],即提出了miRNAs可靶作用调控一些蛋白,亦可被一些蛋白反馈调控的观点,这种机制的复杂性和精密性无疑为神经系统的完整发育提供极大保证。
3 miRNAs表达异常在神经管畸形中的可能作用机制
miRNAs的异常表达可能会对神经管发育做出错误调控,诱使NTD或其它神经系统疾病的发生。Zhang等[33]对孕期14~22周(脑与脊髓发育形成期)的人类无脑儿胚胎进行研究发现,miR-126、miR-198及 miR-451表达量水平与正常胎儿组相比明显上调,而miR-9、-212、-124、-138及miR-103/107则表现为下调,进一步生物信息学的功能分析,约有881个可能的靶基因受这些miRNAs的作用调控,其中79个可能涉及同一蛋白网络的互相作用,据此推断,miRNAs的失调可能是产生无脑儿的致病缘由之一,且是由多个miRNAs表达异常诱发的。有人对RA致畸小鼠的研究发现,GD-13.5、-15.5、-17.5及GD-19.5四期的脊柱裂小鼠胚胎中miRNAs的表达情况具有明显不同,从GD-13.5 到GD-19.5,miR-9/9*、miR-124a和miR-125b的表达量与正常组相比均下降[34],提示这种miRNAs的失调在脊髓的畸化过程中可能是致病因素之一。
miR-34高表达时,它会靶作用于德尔塔样配体1(delta-like-ligand 1,DLL1),从而使Notch蛋白的胞内区不能释放使得Notch通路受阻,引起下游Hes-1、-3、-5蛋白对原神经基因(neurogenin,Ngn)的抑制作用减弱,Ngn蛋白表达量升高,从而影响中-后脑神经发生的形成及dl2中间神经元的建立。而Ngn蛋白这种表达水平的升高,也是过表达的miR-221、-222负调控DNMT1使体内甲基化水平下降的间接结果,即miR-34和miR-221、-222在调节Notch通路中Ngn蛋白的表达水平上表现出一种协同机制。同时,miR-34不仅可直接靶作用于亚甲基四氢叶酸还原酶(methylene tetrahydrofolate reductase,MTHFR),还可通过靶作用于转录因子E2F3间接与miR-24协同沉默二氢叶酸还原酶(dihydrofolate reductase,DHFR),共同对体内一碳代谢(one-carbon metabolism,OCM)和DNA甲基化水平做出负调控,正是miR-34异常高表达时对Notch及DNA甲基化两通路的破坏,才可能诱使细胞过度增殖或分化过早熟导致NTD的发生[35]。
miR-19a及miR-19b可特异靶作用于低密度脂蛋白受体相关蛋白2(low-density lipoprotein receptor-related protein 2,LRP2),已知该蛋白在孕中期小鼠胚胎中的神经上皮组织里高表达,可参与Wnt信号的转导,若定向缺失LRP2基因,则会诱发胚胎死亡[36]。除此之外,miR-19还可靶作用于RhoB(调节背部神经管向神经嵴的层离)[37]、qkl(一旦缺失会导致神经管暴露)[38]、Id2(在颅神经褶中过量表达或在神经嵴迁移会导致神经管发育的早熟)等等[39],若其失调错表达,可能会诱使NTD的发生。
表1是对哺乳动物如小鼠和人类的神经组织中,检测有表达的一些miRNAs及其对神经管发育影响的概括性总结。
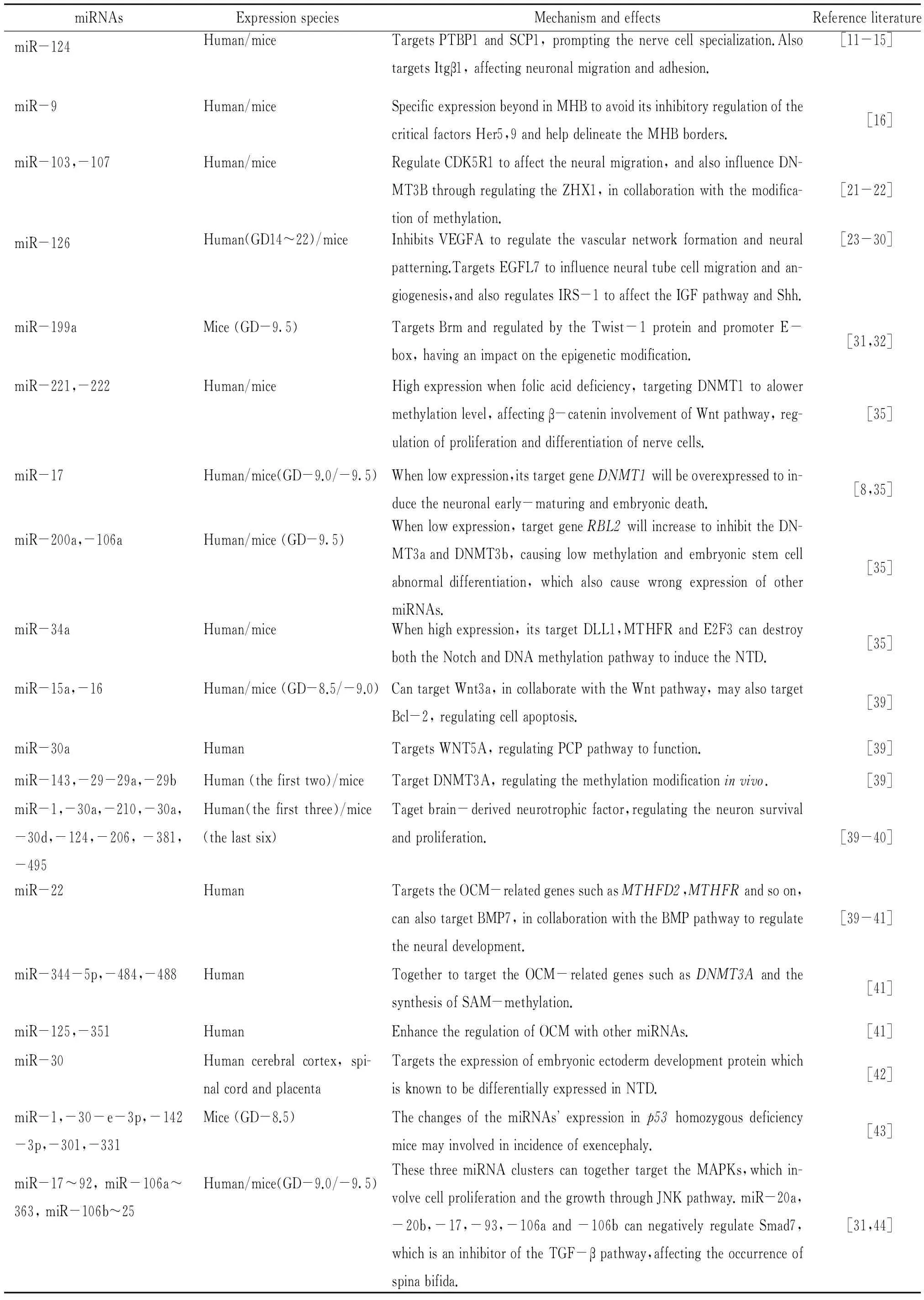
表1 哺乳动物神经组织中检测有表达的miRNAs及其对神经管发育的影响
由该表可以看出,miRNAs在神经系统发育过程中的作用以及表达变异对NTD发病机制的贡献,主要是通过调控Wnt、Notch、细胞凋亡等通路的靶基因来实现的。
4 结论
本文重点阐述了miRNAs在神经管发育期间的作用,总体而言,神经管的正常发育是与其相关基因精确的时空特异性表达,信号转导通路及基因表达调控网络三方面的完美合作是密不可分的,其中任意一方发生变异,都可能诱使NTD的发生。而miRNAs正是在基因的转录后调控中起着不可替代的作用,一些miRNAs阶段特异性的表达失调或对靶基因mRNAs的不准确沉默,都有可能是引起NTD的诱因,但对这方面的研究尚不明确,相信随着科学技术的发展,miRNA作为分子诊断的标志物在NTD中的应用指日可待。
[1] 胡 婷,钟雪云.MicroRNA在脑胶质瘤中的作用[J].中国病理生理杂志,2011,27(7):1431-1437.
[2] 李育敏,费 嘉.MicroRNA与血液系统肿瘤发生、诊断和治疗[J].中国病理生理杂志,2009,25(11):2225-2229.
[3] Copp AJ, Greene ND, Murdoch JN. The genetic basis of mammalian neurulation[J]. Nat Rev Genet, 2003,4(10):784-793.
[4] Greene ND, Copp AJ. Development of the vertebrate central nervous system: formation of the neural tube[J]. Prenat Diagn,2009,29(4):303-311.
[5] Copp AJ, Greene ND.Genetics and development of neural tube defects[J]. J Pathol,2010,220(2): 217-230.
[6] Beaudin AE, Abarinov EV, Malysheva O,et al. Dietary folate, but not choline, modifies neural tube defect risk inShmt1 knockout mice[J].Am J Clin Nutr,2012,95(1):109-114.
[7] Sempere LF, Freemantle S, Pitha-Rowe I, et al. Expression profiling of mammalian microRNAs uncovers a subset of brain-expressed microRNAs with possible roles in murine and human neuronal differentiation[J] . Genome Biol, 2004,5(3):R13.
[8] Mukhopadhyay P, Brock G, Appana S, et al.MicroRNA gene expression signatures in the developing neural tube[J]. Birth Defects Res A Clin Mol Teratol, 2011, 91(8): 744-762.
[9] Deo M, Yu JY, Chung KH, et al. Detection of mammalian microRNA expression byinsituhybridization with RNA oligonucleotides[J].Dev Dyn, 2006, 235(9): 2538-2548.
[10]Cao X, Pfaff SL, Gage FH. A functional study of miR-124 in the developing neural tube[J]. Genes Dev,2007,21(5):531-536.
[11]Makeyev EV, Zhang J, Carrasco MA, et al. The microRNA miR-124 promotes neuronal differentiation by triggering brain-specific alternative pre-mRNA splicing[J]. Mol Cell,2007,27(3):435-448.
[12]Visvanathan J, Lee S, Lee B, et al.The microRNA miR-124 antagonizes the anti-neural REST/SCP1 pathway during embryonic CNS development[J].Genes Dev, 2007,21(7):744-749.
[13]Conaco C, Otto S, Han JJ,et al.Reciprocal actions of REST and a microRNA promote neuronal identity[J].Proc Natl Acad Sci U S A,2006,103(7):2422-2427.
[14]Fournier-Thibault C, Blavet C, Jarov A, et al.Sonic hedgehog regulates integrin activity, cadherin contacts, and cell polarity to orchestrate neural tube morphogenesis[J]. J Neurosci,2009,29(40):12506-12520.
[15]Nagy N, Mwizerwa O, Yaniv K, et al.Endothelial cells promote migration and proliferation of enteric neural crest cells via β1 integrin signaling[J]. Dev Biol, 2009,330(2):263-272.
[16]Leucht C, Stigloher C, Wizenmann A, et al. MicroRNA-9 directs late organizer activity of the midbrain-hindbrain boundary[J]. Nat Neurosci,2008,11(6): 641-648.
[17]Yoo AS, Staahl BT, Chen L, et al. MicroRNA-mediated switching of chromatin remodelling complexes in neural development[J]. Nature,2009,460(7255):642-646.
[18]Krichevsky AM, Sonntag KC, Isacson O, et al.Specific microRNAs modulate embryonic stem cell-derived neurogenesis[J].Stem Cells,2006,24(4):857-864.
[19]Trovato M, D’Armiento M, Lavra L, et al. Expression of p53/ HGF /c- met/STAT3 signal in fetuses with neural tube defects[J]. Virchows Arch,2007,450(2): 203-210.
[20]Babak T, Zhang W, Morris Q, et al.Probing microRNAs with microarrays: tissue specificity and functional inference[J]. RNA,2004,10(11):1813-1819.
[21]Moncini S, Salvi A, Zuccotti P, et al.The role of miR-103 and miR-107 in regulation of CDK5R1 expression and in cellular migration[J].PLoS One,2011, 6(5):e20038.
[22]Kim VN,Nam JW. Genomics of microRNA[J]. Trends Genet,2006,22(3): 165-173.
[23]James JM, Gewolb C, Bautch VL.Neurovascular development uses VEGF-A signaling to regulate blood vessel ingression into the neural tube[J]. Development, 2009,136(5):833-841.
[24]Ie Roux I, Lewis J, Ish-Horowicz D.Notch activity is required to maintain floorplate identity and to control neurogenesis in the chick hindbrain and spinal cord[J]. Int J Dev Biol,2003,47(4):263-272.
[25]Sun Y, Bai Y, Zhang F, et al.MiR-126 inhibits non-small cell lung cancer cells proliferation by targeting EGFL7[J]. Biochem Biophys Res Commun,2010, 391(3): 1483-1489.
[26]Parathath SR, Mainwaring LA, Fernandez LA, et al.Insulin receptor substrate 1 is an effector of sonic hedgehog mitogenic signaling in cerebellar neural precursors [J]. Development,2008,135(19):3291-3300.
[27]Bernd P.The role of neurotrophins during early development[J]. Gene Expr, 2008,14(4):241-250.
[28]Zhang J, Du YY, Lin YF, et al.The cell growth suppressor, mir-126, targets IRS-1[J]. Biochem Biophys Res Commun,2008,377(1):136-140.
[29]Shen WF, Hu YL, Uttarwar L, et al.MicroRNA-126 regulates HOXA9 by binding to the homeobox[J]. Mol Cell Biol,2008,28(14):4609-4619.
[30]Whelan JT, Ludwig DL, Bertrand FE.HoxA9 induces insulin-like growth factor-1 receptor expression in B-lineage acute lymphoblastic leukemia[J]. Leukemia, 2008,22(6):1161-1169.
[31]Sakurai K, Furukawa C, Haraguchi T, et al. microRNAs miR-199a-5p and -3p target the Brm subunit of SWI/SNF to generate a double-negative feedback loop in a variety of human cancers[J]. Cancer Res,2011,71(5):1680-1689.
[32]Lee YB, Bantounas I, Lee DY, et al. Twist-1 regulates the miR-199a/214 cluster during development[J]. Nucleic Acids Res,2009,37(1):123-128.
[33]Zhang Z, Chang H, Li Y, et al. MicroRNAs: potential regulators involved in human anencephaly[J]. Int J Biochem Cell Biol,2010,42(2):367-374.
[34]Zhao JJ, Sun DG, Wang J, et al. Retinoic acid downregulates microRNAs to induce abnormal development of spinal cord in spina bifida rat model[J]. Childs Nerv Syst,2008,24(4):485-492.
[35]Shookhoff JM,Gallicano GI.A new perspective on neural tube defects: folic acid and microRNA misexpression[J].Genesis,2010,48(5):282-294.
[36]Willnow TE, Hilpert J, Armstrong SA, et al.Defective forebrain development in mice lacking gp330 / megalin[J].Proc Natl Acad Sci U S A ,1996,93(16): 8460-8464.
[37]Wang Y, Stricker HM, Gou D, et al.MicroRNA: past and present[J].Front Biosci,2007,12: 2316-2329.
[38]Li Z, Takakura N, Oike Y, et al.Defective smooth muscle development in qkI-deficient mice[J]. Dev Growth Differ,2003,45(5-6):449-462.
[39]Hsu SD, Lin FM, Wu WY,et al.miRTarBase: a database curates experimentally validated microRNA-target interactions[J].Nucleic Acids Res,2011,39(Suppl 1):D163-D169.
[40]Jungbluth S, Koentges G, Lumsden A. Coordination of early neural tube development by BDNF/trkB[J]. Development,1997,124(10):1877-1885.
[41]Stone N, Pangilinan F, Molloy AM,et al.Bioinformatic and genetic association analysis of microRNA target sites in one-carbon metabolism genes[J]. PLoS One,2011,6(7): e21851.
[42]Song PP, Hu Y, Liu CM, et al. Embryonic ectoderm development protein is regulated by microRNAs in human neural tube defects[J].Am J Obstet Gynecol,2011, 204(6):544.e9-544.e17.
[43]Hosako H,Martin GS,Barrier M,et al. Gene and microRNA expression inp53-deficient day 8.5 mouse embryos[J]. Birth Defects Res A Clin Mol Teratol, 2009, 85(6):546-555.
[44]Nakayama T, Berg LK, Christian JL.Dissection of inhibitory Smad proteins: both N- and C-terminal domains are necessary for full activities ofXenopusSmad6 and Smad7[J]. Mech Dev,2001,100(2):251-262.
TemporalandspatioaleffectsofenrichedmiRNAsinbrainandpossibleroleinneuraltubedefects
MicroRNAs (miRNAs) are known to play an important role in regulating the cell proliferation, differentiation and organogenesis. Abnormal expression of miRNAs is associated with increased risk for various diseases, such as neural tube defects (NTD), which is a severe developmental disease as the result of closing the neural tube incompletely or too early. The present review summarizes some enriched miRNAs in the brain of mammals and their spatial temporal feature, and tries to elaborate the relationship between the aberrant expression of miRNAs and the pathogenesis of NTD.
微小RNA; 神经管缺陷; 哺乳动物
MicroRNA; Neural tube defects; Mammals
BAI Bao-ling, LI Hui-li, CHEN Shu-yuan, WANG Xiao-lei, ZHANG Qin, ZHAO Hui-zhi, ZHANG Ting
(CapitalInstituteofPediatrics,Beijing100020,China.E-mail:zhangtingcv@yahoo.com.cn)
R394.3
A
10.3969/j.issn.1000-4718.2012.12.034
