三叶鬼针草生长特征对土壤氮素水平的响应
2012-11-06潘玉梅唐赛春韦春强刘明超
潘玉梅, 唐赛春, 韦春强, 刘明超,2
(1.广西壮族自治区中国科学院广西植物研究所,广西桂林 541006; 2.广西师范大学,广西桂林 541004)
潘玉梅,唐赛春,韦春强,等. 三叶鬼针草生长特征对土壤氮素水平的响应[J]. 杂草科学,2012,30(1):11-16.
三叶鬼针草生长特征对土壤氮素水平的响应
潘玉梅1, 唐赛春1, 韦春强1, 刘明超1,2
(1.广西壮族自治区中国科学院广西植物研究所,广西桂林 541006; 2.广西师范大学,广西桂林 541004)
利用温室盆栽试验法,研究入侵植物三叶鬼针草生长特征对不同土壤氮素水平的响应。结果表明,氮素的增加能够促进三叶鬼针草叶片数、总叶面积、分枝数及生物量的增加,提高其相对生长速率。在物质分配方面,氮肥的施用能促进该植物向地上部分分配较多的物质,显著增加生殖器官(花)生物量的投入,降低向地下部分(根)的物质投入。同时,可塑性指数表明,三叶鬼针草在形态、生物量分配及生殖器官等方面对土壤氮素响应的表型可塑性较大。以上结果充分体现了三叶鬼针草具有较强的入侵性和对土壤氮素环境较高的适应性,预警人们对其预防和管理工作应加以重视。
三叶鬼针草; 表型可塑性; 入侵性; 氮素
近年来,在全球环境变化问题研究大背景下,外来生物入侵带来的一系列生态环境问题成为生态学研究的焦点之一[1-2]。弄清生物的入侵性和环境的可入侵性是研究生物入侵机制的两个关键因素[3-4]。外来植物的入侵性不仅与自身生长力、繁殖特性有关,还与入侵地的生态环境因子及其对入侵地异质性环境的适应能力密切相关[5]。因此,研究外来入侵植物生长特征与外界生态环境因子的关系,对弄清外来植物入侵性的主要影响因素,探讨外来植物的入侵机理有重要意义。
土壤养分是限制植物生长的重要环境因子之一。有研究表明,高养分环境中外来入侵植物具有较高的生物量[6-7]、较大的相对生长速率[8]和较高的养分利用效率[9]。在贫瘠的环境中,养分的增加能促进环境的被入侵性,如土壤磷素的增加促进了澳大利亚草地生态系统的外来种入侵[10],且生境养分的水平还直接影响着入侵种的入侵程度[11]。氮肥是影响植物生长最重要的土壤养分之一,影响着植物的生长和生物量分配。研究表明,在一定范围内,植物的叶面积比随着土壤有效氮的增加而增加,根冠比则随着有效氮的增加而降低[12-13]。研究入侵植物对土壤氮素水平的响应,一方面可以反映出该植物对土壤养分的适应能力,另一方面根据氮素与入侵植物入侵性的关系,可以初步预测该植物可能的入侵范围,为入侵植物的预防和管理提供依据。
我国外来入侵植物中,菊科外来物种占据较大的比例。三叶鬼针草(BidenspilosaL.)为菊科(Compositae)鬼针草属(BidensL.)一年生草本植物,在生物学特性方面,该物种不仅具有菊科外来物种的共性,如生长发育快、果实量大、成熟早等特性[14],还具有其特殊的生物学特性,如花果期较长,可以不断地产生种子,萌发率高,繁殖力强、瘦果冠毛芒状且具倒刺,易被携带而传播。该物种原产热带美洲,现广泛分布于亚洲和美洲的热带、亚热带地区[15],已入侵我国多个省区,尤其在西南、华南地区入侵严重,是农业、林业、畜牧业等主要的有害外来植物之一,严重威胁到当地生物多样性,破坏生态平衡,被多个省区归化为入侵性较强的入侵种[16-17]。由于其目前主要入侵公路、农田和林下局部区域等人为干扰较大的生境,入侵程度还未像紫茎泽兰(EupatoriumadenophorumSpreng)、飞机草(EupatoriumodoratumLinn.)、水葫芦(Eichhorniacrassipes)等那么严重,仅在局部地区造成严重危害,常被人们忽视,因此,对其入侵性的相关研究较少。已有的研究主要关于其分布与危害状况[18]、化感特性[19,20]、种子萌发[21]及其繁殖特征与入侵性的关系[22]等,关于其入侵性与环境因子之间关系的研究仅见王瑞龙等[23]报道了三叶鬼针草化感特性与大气环境温度的关系。本研究通过盆栽试验法研究三叶鬼针草形态、生物量、生物量分配及繁殖结构等对土壤氮素水平的响应,为弄清影响该植物入侵性的环境因素,探讨其入侵机理有重要作用。
1 材料与方法
试验在广西植物研究所温室大棚内进行,地理位置为110°12′E,25°11′N,海拔170 m,属于中亚热带季风气候区,全年平均温度19.2 ℃,年降雨量1 655.6 mm。于5月初将籽粒饱满的三叶鬼针草种子均匀播种在花盆中育苗,待幼苗长至高10 cm左右,选取长势一致的幼苗移栽到内径为23 cm,深为18 cm的盆中,每盆1株,栽培基质由肥土与砖红壤等体积混合而成,每盆装土3 kg,经广西农业科学院土壤肥料研究所测定,土壤基质pH值为7.65,含有机质21.58 g/kg,全N 0.096 g/kg,全P 0.074 g/kg,全K 0.975 g/kg,有效N 91 mg/kg,有效P 17 mg/kg,有效K 65 mg/kg。在遮阳条件下进行为期两周的适应性生长。于6月底对盆栽的三叶鬼针草进行土壤氮养分含量的处理。设置为:CK(不施氮)、N1(0.1 g/kg)、N2(0.2 g/kg)、N3(0.4 g/kg)4个梯度,5次重复。施肥方法是先定量称取肥料,用水稀释后分3次施入,每次间隔为10 d。施肥前随机选取5~8株幼苗测定其初始生物量和初始总叶面积,以便进行生长速率、净同化速率等生长特性的计算。所选氮肥为尿素(含有效N 46%),试验期间每天傍晚浇足水,随时防治病虫害。
待植株处于营养生长旺盛期,测量其形态生长参数,即株高、总叶面积、叶片数及总生物量、根生物量、叶生物量、茎生物量、分枝数等。用Li-3000 型叶面积仪测量叶面积,称重部分在80 ℃下烘48 h,电子天平(精确度0.000 1 g)称量。计算如下参数:叶面积比(总叶面积/植株总重),比叶面积(总叶面积/总叶重),根生物量比(根重/植株总重)、叶生物量比(叶重/植株总重)、支持结构生物量比(支持结构重/植株总重)、根冠比(根生物量/地上部分生物量)。采用Poorter[24](1999)的方法计算平均相对生长速率(RGR)、净同化速率(NAR)和平均叶面积比LARm。公式如下:
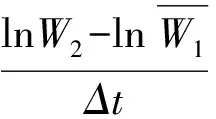
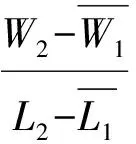
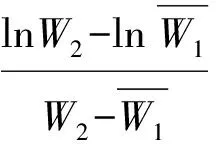
待植株处于繁殖期,测量头状花序数、种子干重,计算种子千粒重。利用SPSS软件的One-way ANOVA方法对数据进行处理和分析,利用Excel软件绘图。
2 结果与分析
2.1 土壤氮素水平对三叶鬼针草形态特征的影响
对不同氮素处理水平下三叶鬼针草的形态特征进行比较,结果发现氮素处理对植株株高无显著影响,各处理的植株株高差异不明显(P>0.05)。施氮处理对植株叶片数、总叶面积、分枝数、叶面积比和比叶面积均有较大影响,表现为叶片数、总叶面积及分枝数随着氮素水平的增加而显著增加(P<0.05)。叶面积比和比叶面积在高氮素处理下显著高于中、低氮处理和对照(P<0.05,图1)。说明氮素的增加主要促进三叶鬼针草叶和分枝的增长,各形态参数的可塑性指数中叶片数、叶面积及分枝数的可塑性指数较高也说明了这点(表1)。
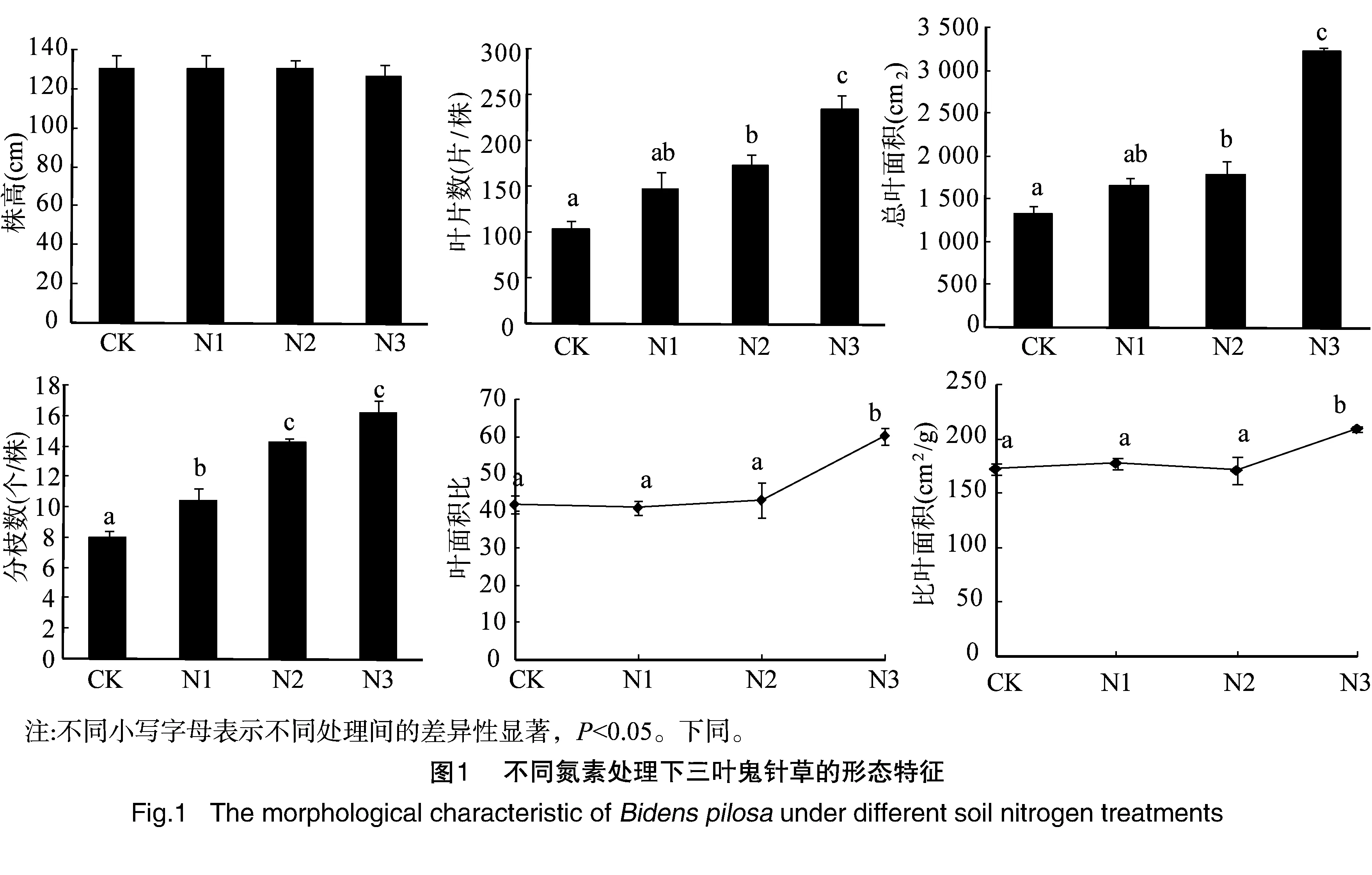
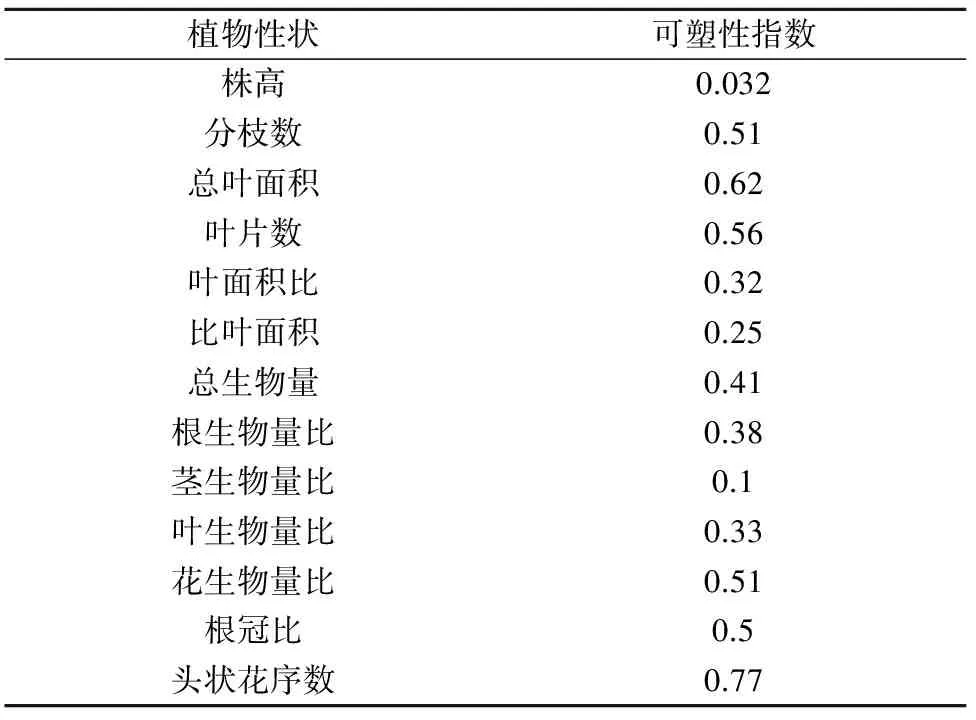
表1 三叶鬼针草形态、生物量及物质分配对氮素水平响应的表型可塑性指数
2.2 土壤氮素水平对三叶鬼针草生物量积累及物质分配的影响
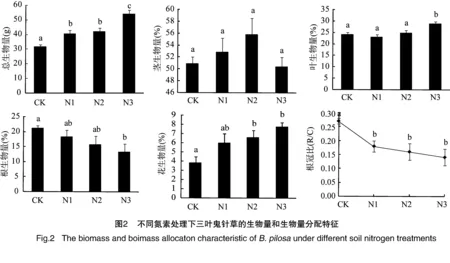
对各处理下三叶鬼针草的总生物量及物质分配进行了分析,结果显示施氮处理对植株总生物量影响显著,随着施氮量的增加而显著增加(P<0.05)。对茎生物量比无明显影响(P>0.05)。叶生物量比在高氮处理下显著高于中、低氮处理和对照(P<0.05)。但中、低氮处理和对照间无显著差异(P>0.05)。根生物量比和根冠比随着施氮量增加而减小,高氮处理下显著低于对照(P<0.05)。花生物量比随着施氮量增加而增加,与对照相比,中、高氮处理增加的较显著(P<0.05)。可见一定量的氮素能增加三叶鬼针草植株的总生物量,促进物质向地上部分分配,而减少对地下部分的物质投入(图2)。
2.3 土壤氮素水平对三叶鬼针草生长特性及繁殖特征的影响
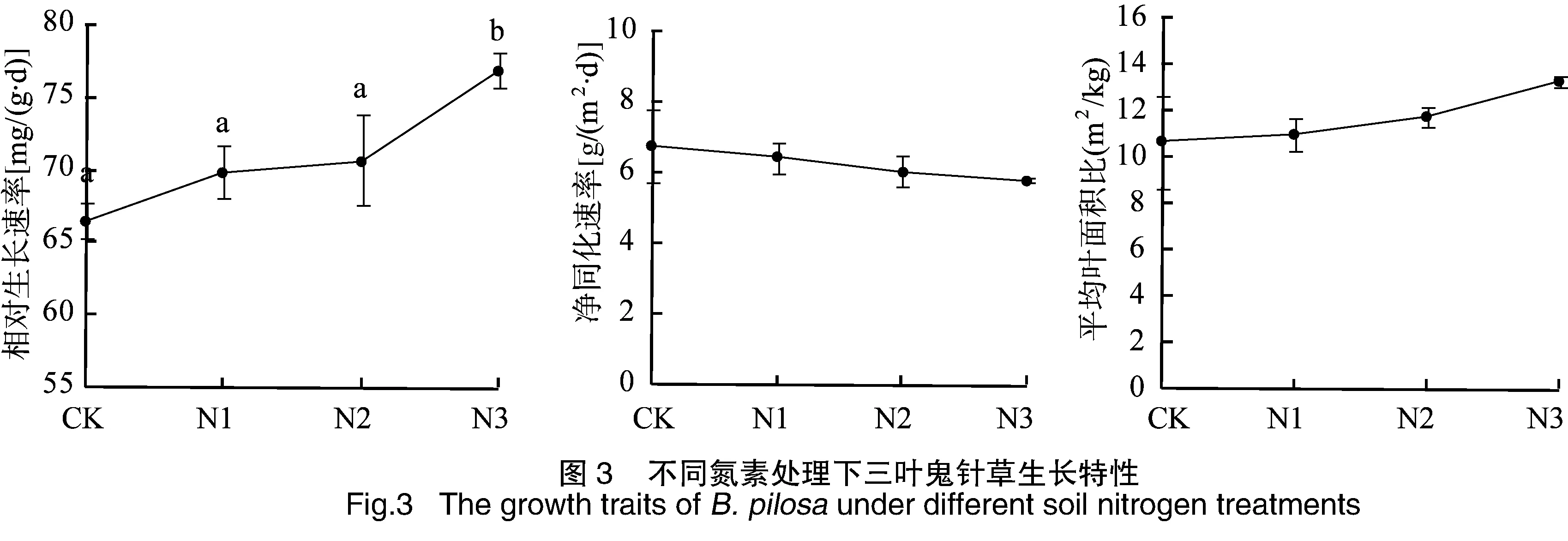
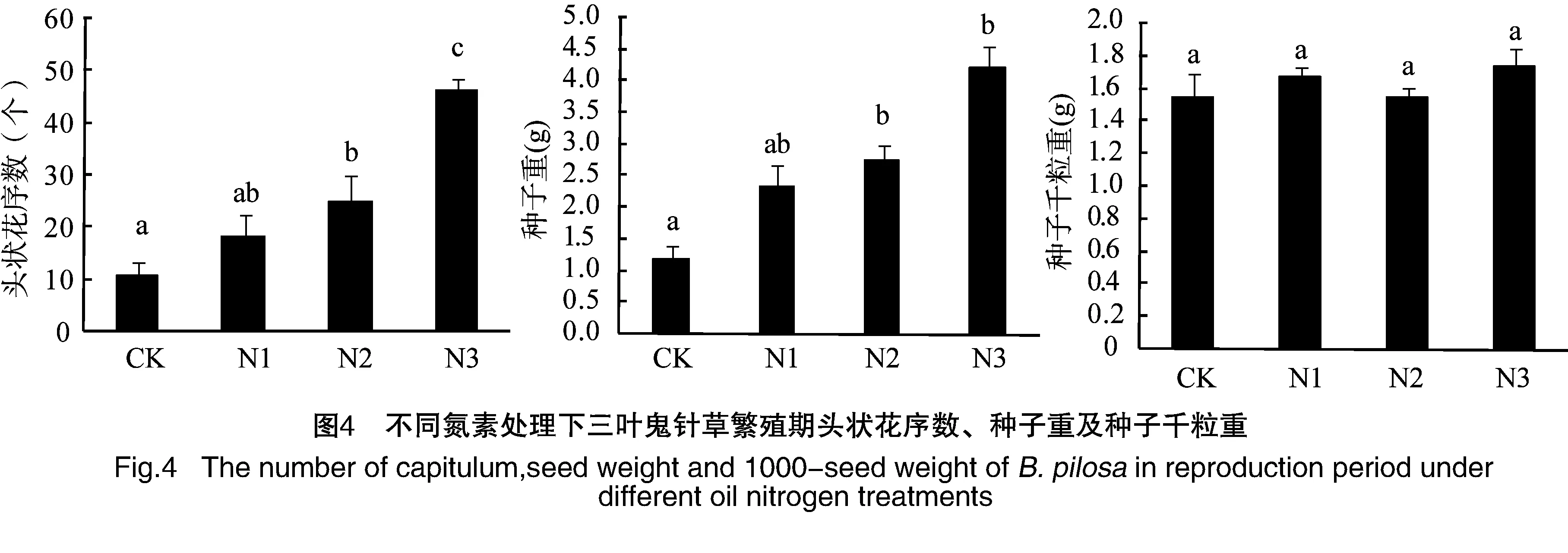
对不同氮素处理下三叶鬼针草生长旺盛期的生长特性及繁殖特征进行了比较,三叶鬼针草的相对生长速率和平均叶面积比均随着施氮量增加而逐渐增加,高氮处理下相对生长速率明显高于其他氮素处理,而平均叶面积比增加的均不明显。净同化速率随着施氮量增加而减小,但减小的不明显(图3)。三叶鬼针草头状花序数和种子干重随着施氮量增加而显著增加(P<0.05),而种子千粒重在各氮素处理间无明显差异(P>0.05,图4)。说明氮素达到一定程度时能显著增加三叶鬼针草的生长速率,同时也能促进繁殖结构的生长,如花的数量和结实量的增加等,头状花序数和花生物量比的可塑性指数较大,也说明了三叶鬼针草繁殖结构对氮养分响应的表型可塑性较大(表1)。
3 讨论
对外来入侵植物的预测及监控的关键是在外来植物入侵之前评价其入侵力[25],而评价外来植物入侵力的主要任务之一就是弄清影响外来植物成功入侵的影响因素[26]。研究表明,养分的增加可促进外来种的入侵[10],高养分环境中入侵植物竞争力更强,从而进一步加剧入侵[27-28]。本研究中施氮量的增加可以明显提高三叶鬼针草的总生物量、花生物量比和相对生长速率,说明土壤环境中氮肥的增加在一定程度上可以促进三叶鬼针草的入侵,与以上学者们的研究结论一致。同时,在高氮素处理下,三叶鬼针草根生物量比和根冠比显著降低,叶片数、总叶面积和分枝数显著增加。当土壤养分充足时,减少对地下部分物质的分配,适当的增加地上部分物质的投入,尤其增加光合结构和分枝数的投入,可以更好的竞争地上部分可利用的光和空间等资源,这是三叶鬼针草对高养分环境做出的自我生长调节策略,可能是其在高养分环境中维持较强生长竞争力的主要策略之一,体现了该植物对高氮养分环境的适应性。
表型可塑性是同一基因型应对不同环境条件产生不同表型的特性,是生物在没有遗传变异的情况下适应环境的一种机制。在异质生境中,表型可塑性强的植物能通过改变其形态、生长、生物量分配等增加对各种资源的获得、占据多样的生境[29]。表型可塑性与外来物种入侵能力呈正相关关系[30],具有较大表型可塑性也是成功入侵植物的重要特征之一[31]。本研究中三叶鬼针草的形态参数和生物量分配参数如分枝数、总叶面积、叶片数、花生物量比等的表型可塑性指数均相对较高,表明三叶鬼针草对氮素响应的表型可塑性较大。较大的表型可塑性使三叶鬼针草在氮肥贫瘠的土壤环境中减少对光合结构叶的投入,降低光合作用,增加对地下根生物量的投入,利用根的吸收功能竞争地下营养资源,以维持植株较好的生长。而在氮肥充足的土壤环境中增加地上部分的物质投入,通过增加叶片数和总叶面积加强光合作用,加大竞争和充分利用光能的能力,适当的降低了对地下部分物质的投入。体现出三叶鬼针草与其他成功入侵植物的共性。
繁殖是入侵种群建群时面临的根本挑战,与繁殖有关的特征直接影响到归化和入侵的成功[32,22]。入侵植物的繁殖能力与入侵能力呈正相关[33-34]。郝建华等(2009)研究认为三叶鬼针草结实量大以及种子萌发范围广、易传播等繁殖特征是增强其入侵性的重要因素之一。本研究中施氮量的增加显著提高了三叶鬼针草的开花数和种子干重,表明氮肥促进了三叶鬼针草向繁殖结构物质的投入。同时,不同氮素处理下头状花序数的表型可塑性指数达到了0.77,高于其他参数,说明三叶鬼针草的繁殖结构在异质土壤养分环境中的可塑性比其他部位的更高,揭示出三叶鬼针草的繁殖结构对该植物入侵成功的重要作用。对三叶鬼针草与本地同属近缘种的比较研究中也肯定了繁殖结构在三叶鬼针草与本地种竞争中能够占据优势的意义。同属物种之间的比较研究表明,结实性、有效的扩散模式、萌发能力以及幼苗延存和建群能力等繁殖特征,是促进植物入侵性的因素[33],三叶鬼针草在我国南方入侵地具有一年四季开花结实,结实量大,且种子萌发能力强、萌发速度快等繁殖特征,具备了促进植物入侵性的繁殖特性。因此,三叶鬼针草这种繁殖特性可以认为是该植物成功入侵的一个因素,而氮肥的增加能够促进该植物繁殖结构的生长。由此断定,土壤环境中氮素的多寡是影响该植物入侵成功的重要因素之一。
4 小结
三叶鬼针草对不同氮素的处理,在形态、生物量分配及繁殖特征方面表现出较大的表型可塑性,体现出对土壤高氮环境的适应性。氮肥能够促进该植物的入侵,增强其入侵性。因此,在对该植物进行预防和管理时,应尽量避免将该植物丢弃在含氮较高的环境中,以防其残留种子萌发从而大量繁殖。另外,在人为干扰的生境中,尽量减少垃圾的堆放,以避免为该植物创造高养分环境,减少促进其入侵的外界因素。尽管目前三叶鬼针草仅对局部区域造成严重危害,还未引起人们高度重视,但其较强的入侵性及较高的繁殖力,一旦得到有利的环境,其入侵势头将不可抵挡。因此,有必要对该植物的影响因素及其入侵性进行研究,也为以后该植物的控制和管理提供基础依据。
[1]万方浩,郭建英,王德辉. 中国外来生物的危害与管理对策[J]. 生物多样性,2002,10(1):119-125.
[2]Strong D R,Pemberton R W. Biological control of invading species-risk and reform [J]. Science,2000,288:1969-1970.
[3]Gerlach J D,Rice K J. Testing life history correlates of invasiveness using congeneric plant species[J]. Ecological Applications,2003,13:167-179.
[4]Leicht-Young S A,Silander J A Jr,Latimer A M. Comparative performance of invasive and nativeCelastrusspecies across environmental gradients[J]. Oecologia,2007,154:273-282.
[5]Ehrenfeld J G.Effects of exotic plant invations on soil nutrient cycling processes[J].Ecosystems,2003,36:503-523.
[6]王 坤,杨 继,陈家宽. 不同土壤水分和养分条件下喜旱莲子草与同属种生长状况的比较研究[J]. 生物多样性,2010,18(6):615-621.
[7]Hwang B C,Laueroth W K. Effect of nitrogen,water and neighbor density on the growth ofHesperismatronalisand two native perennials[J]. Biological Invasions,2008,10(5):771-779.
[8]王满莲,冯玉龙,李 新. 紫茎泽兰和飞机草的形态和光合特性对磷营养的响应[J]. 应用生态学报,2006,17(4):602-606.
[9]Zou J W,Rogers W E,DeWalt S J,et al. The effect of Chinese tallow tree(Sapiumsebiferum)ecotype on soil-plant system carbon and nitrogen pro-cesses[J]. Oecologia,2006,150:272-281.
[10]Scott A K,Buckney R T. Invasion of exotic plants in nutrient-enriched urban bushland[J]. Aust Eco1,2002,27:573-583.
[11]Cale P,Hobbs R J. Condition of Zorro:Effects of the exotic annual grassVulpiamyuroson California native perennial grasses [J]. Restor Ecol,1991,8:l0-l7.
[12]Meziane D,Shipley B. Interacting determinants of specific leaf area in 22 herbaceous species:effects of irradiance and nutrient availability [J]. Plant Cell and Environment,1999,22(5):447-459.
[13]Elberse I A M,van Damme J M M,van Tienderen P H. Plasticity of growth characteristics in wild barley(Hordeumspontaneum) in response to nutrient limitation[J]. Journal of Ecology,2003,91:371-382.
[14]朱世新,覃海宁,陈艺林.中国菊科植物外来种概述[J]. 广西植物,2005,25(1):69-76.
[15]李振宇,解 炎.中国外来入侵种[M]. 北京:中国林业出版社,2002:158.
[16]徐海根,强胜.中国外来入侵物种编目 [M]. 北京:中国环境科学出版社,2004:407-409
[17]万方浩,郑小波,郭建英.重大农林外来入侵物种的生物学与控制[M]. 北京:科学出版社,2005:774.
[18]周国庆,官 旋,彭友林,等. 常德市外来物种三叶鬼针草的形态建成与危害研究 [J]. 安徽农业科学,2010,38(2):880-882.
[19]杜凤移,张苗苗,马丹炜,等.三叶鬼针草化感作用的初步研究[J]. 中国植保导刊,2007,27(9):8-11.
[20]贾海江,唐赛春,李先琨,等. 三叶鬼针草对岩溶木本植物任豆和香椿的化感作用[J],广西科学,2008,15(4):436-440.
[21]洪 岚,沈 浩,杨期和,等.外来人侵植物三叶鬼针草种子萌发与贮藏特性研究[J]. 武汉植物学研究,2004,22(5):433-437.
[22]郝建华,刘倩倩,强 胜. 菊科入侵植物三叶鬼针草的繁殖特征及其与入侵性的关系 [J]. 植物学报,2009,44(6):656-665.
[23]王瑞龙,韩 萌,梁笑婷,等.三叶鬼针草生物量分配和化感作用对大气环境温度升高的响应[J]. 生态环境学报,2011,20(6-7):1026-1030.
[24]Poorter L.Growth response of 15 rain-forest tree species to a light gradient:the relative importance of morphological and physiological traits [J].Functional Ecology,1999,13:396-410.
[25]Goodwin B J,McAllister A J,Fahrig L. Predicting invasiveness of plant species based on biological information[J]. Conservation Biology,1999,13:422-426.
[26]Heger T,Trepl L. Predicting biological invasions [J]. Biological Invasions,2003,5:313-321
[27]Brooks M L. Effects of increased soil nitrogen on the dominance of alien annual plants in the Mojave Desert[J]. Journal of Applied Ecology,2003,40:344-353.
[28]田耀华,冯玉龙,刘朝.氮肥和种植密度对紫茎泽兰生长和竞争的影响[J]. 生态学杂志,2009,28(4):577-588.
[29]张常隆,李扬苹,冯玉龙,等. 表型可塑性和局域适应在紫茎泽兰入侵不同海拔生境中的作用[J]. 生态学报,2009,29(4):1940-1946.
[30]耿宇鹏,张文驹,李 博,等. 表型可塑性与外来植物的入侵能力[J]. 生物多样性,2004,12(4):447-455.
[31]Baker H G. Characteristics and models of origin of weeds[M]//Baker H G,Stebbins G L. The genetics of colonizing species. NewYork:Academic Press,1965:147-169.
[32]Pyšek P,Richardson D M. Traits associated with invasiveness in alien plants:Where do we stand? [M]//Nentwig W. Biological Invasions:Section II. Berlin:Verlag-Springer,2007:97-125.
[33]Barret S C H. Cropmimicry in weeds[J]. Econ Bot,2007,37:255-282.
[34]徐汝梅,叶万辉. 生物入侵理论与实践[M]. 北京:科学出版社,2003:17-18.
ResponseofGrowthCharacteristicsofBidenspiolsaL.toSoilNitrogenLevel
PAN Yu-mei1, TANG Sai-chun1, WEI Chun-qiang1, LIU Ming-chao1,2
(1. Guangxi Institute of Botany,Guangxi Zhuangzu Autonomous Region and Chinese Academy of Sciences,Guilin 541006,China; 2. Guangxi Normal University,Guilin 541004,China)
Soil nitrogen effects on the growth characteristics of alien invasive plantBidenspilosaL. were studied by greenhouse pot experiment. The result indicated that added definite soil nitrogen were favorable to increase the number of leaf and branch,total leaf area(TLA),total biomass and relative growth rate(RGR)ofB.pilosa. In the meantime,increased soil nitrogen could facilitate the weed assign more material to the organs on the ground,especially enhance the flower biomass rate significantly,and decrease the material assigned to the roots under ground. Phenotypic plasticity index indicated that in the case of morphological parameters,biomass allocation parameters and reproduction organ more phenotypic plasticity the weed presented on response to the soil nitrogen. All results above demonstrated that the weedB.pilosahad more grater invasion and adaptability to the soil nitrogen,which suggested that definite attention should be paid to the work of the weed prevention and management.
BidenspilosaL.; phenotypic plasticity; invasiveness; soil nitrogen level
Q142.3;Q142.9
A
1003-935X(2012)01-0011-06
2012-01-11
广西自然科学基金北部湾重大专项 (编号:2011GXNSFE 018005);广西植物研究所科学研究基金(编号:桂植业09028;桂植业10011)。
潘玉梅(1981—),女,安徽萧县人,助理研究员,主要从事外来入侵物种的研究。Email:panyumei@gxib.cn。
唐赛春。Email: tangs@gxib.cn。
