拟南芥和大白菜LUG蛋白家族的生物信息学分析
2012-10-26马宗琪王凤德邱念伟
马宗琪,王凤德,邱念伟
拟南芥和大白菜LUG蛋白家族的生物信息学分析
马宗琪1,王凤德2,*邱念伟1
1.师范大学生命科学学院,山东,曲阜 273165; 2.山东省农业科学院蔬菜研究所,山东,济南 250100)
LUG蛋白属于WD40超家族基因,具有7个重复的WD40结构域,参与花器官的发育。本项研究系统鉴定了7个拟南芥和5个大白菜的LUG基因,并对这些基因编码的蛋白质序列进行了序列保守性和系统进化分析。结果表明,除了AtLUG6含有5个重复的WD40结构域之外,其余所有的LUG蛋白都具有7个重复的WD40结构域。在进化上,LUG蛋白家族可分为两个不同的亚家族,并且这种特征在拟南芥和大白菜分离之前就已经形成。
拟南芥();大白菜();生物信息学分析;LUG蛋白;WD40结构
关于花器官发育的一个重要模型是ABC模型,其基本理论是将花分为萼片、花瓣、雄蕊和雌蕊4轮,这4轮花器官分别由A、B、C三类基因调控,其中A类基因调控萼片的发育,A和B类基因共同作用调控花瓣的发育,B和C类基因共同决定雄蕊的发育,C类基因决定雌蕊的发育[1]。在拟南芥中,A类基因以()、()为代表,B类基因以()和()为代表,C类基因以()为代表。其中C类基因决定内两轮,即雄蕊和雌蕊的发育,只在内两轮中表达。LEUNIG (LUG)是在拟南芥中发现的调节基因表达的转录抑制因子,参与拟南芥的花发育过程[2]。在()和()突变体中,基因异位地在花的外两轮表达,导致萼片转化成心皮,花冠转化成雄蕊,说明基因的功能是在花的外两轮中抑制基因的表达[3-4]。此外,也参与调节其它与无关的发育过程, 例如突变体还表现出柱头裂开、心皮和胚珠发育不正常、雌性和雄性育性降低,花和叶子狭窄等性状[5-7]。
分子生物学的分析表明,LUG属于WD40蛋白家族,在其C-末端含有7个WD重复,2个谷氨酰胺富含区域[2]。已有的研究表明, LUG没有DNA 结合序列,所以LUG与顺式调节原件互作调节的转录很可能还有其它蛋白的参与[8]。此外, LUG与转录因子的互作可能是通过WD重复单元进行的。因为突变表型最严重的是失去最后一个WD重复单元的突变体,说明WD重复对LUG的功能行使是不可或缺的[7]。另外LUG可能在不同的组织中或不同发育阶段与不同的DNA结合因子互作,行使不同的功能,这种表型和表达的广泛性与LUG功能的多样性是一致的[8]。本研究基于已报道的拟南芥家族基因序列为探针对大白菜基因组数据库进行了BLASTN搜索,寻找大白菜相似的候选基因。然后对拟南芥和大白菜候选基因进行了生物信息学分析,包括基因结构、进化关系等。
1 材料与方法
1.1 拟南芥LUG家族基因序列获得及大白菜LUG家族成员搜索
拟南芥家族基因序列下载自DATF网站(http://datf.cbi.pku.edu.cn/search_result.php)。然后以拟南芥基因序列为探针,通过BLASTN程序在大白菜基因组数据库(http://brassicadb.org/ brad/)中搜索相似的基因序列,然后利用DNAMAN软件预测其编码氨基酸序列。将预测的编码氨基酸序列通过BLASTP程序在NCBI数据库进行检测,若属于WD40超家族,并含有7个WD40重复则认为其属于大白菜蛋白家族候选成员。
1.2 基因结构分析、序列比对和系统进化树分析
基因结构分析采用网上在线分析软件GSDS(http://gsds.cbi.pku.edu.cn/);基因的5'UTR区多序列核苷酸比对分析和和LUG蛋白家族的氨基酸多序列比对分析采用DNAMAN软件,采用默认参数设置;拟南芥和大白菜中LUG蛋白家族成员的系统进化分析使用MEGA4.0软件,采用Bootstrap test-Neighbor Joining方法,重复500次运算。
2 结果与分析
2.1 拟南芥和大白菜LUG基因结构和染色体分布分析
根据DATF网站(http://datf.cbi.pku.edu.cn/search _result.php)公布的拟南芥LUG基因序列可以看出,拟南芥基因组共存在7个基因,分别命名为(表1)。其中拟南芥分布于染色体2,位于条染色体4。进一步的分析发现,的编码区都位于相同的染色体位点,由17个相同的外显子组成(图1),其差异仅存在于5’UTR区(图2)。对于基因蛋白编码区,其大部分序列与相同,仅在3’端缺失了最后两个外显子和倒数第三个外显子的部分序列(图1)。因此,推测这6个基因可能是由相同的亲本序列经过不同的RNA编辑形成。
通过Blast搜索和比对,在现有的大白菜基因组数据中鉴定出5个LUG蛋白家族基因,分别命名为(表1)。其中、和分别位于染色体5、1和8,和则位于染色体3。编码区的外显子数目分别为:14、18、25、17和19个(图1)。据此可以看出,与拟南芥相比,大白菜家族成员在染色体上的位点、基因序列结构等方面更为复杂,这可能与家族成员在进化中为了适应和参与更多的生物调节过程有关,也可能与大白菜具有复杂的基因组序列有关。

图1 拟南芥和大白菜LUG基因结构

图2 拟南芥AtLUG1-AtLUG6基因5'UTR区比对
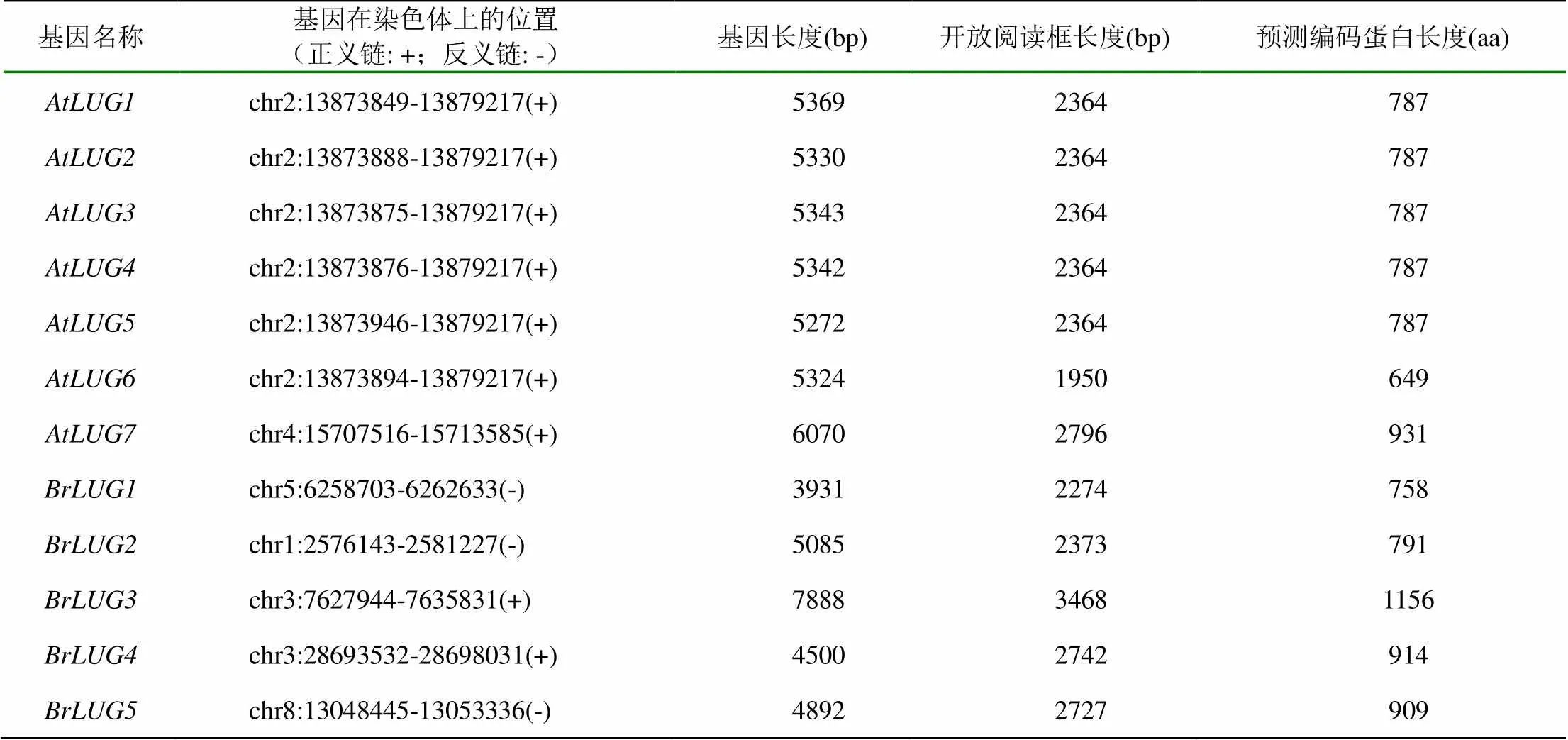
表1 拟南芥和大白菜LUG基因相关信息
2.2 LUG蛋白的多序列联配分析
为了确定植物中LUG蛋白的序列特征,用DNAMAN软件对3个拟南芥(由于具有相同的氨基酸编码序列,所以仅选择AtLUG1进行分析)和5个大白菜的蛋自家族的N端和C端氨基酸进行了多序列比对分析,结果表明供试序列之间存在较高的同源性,在N端除了和含有一个Q-rich区外,其它都含有两个Q-rich区(图 3a)。通过对LUG蛋白的C端进行分析发现,除了由于出现序列缺失具有5个WD40重复外,其它成员都具有7个相对保守的WD40重复(图 3b)。在其C端由于序列缺失所造成的WD40重复的减少是否影响其生物学功能还有待于进一步研究,这对于研究WD40结构域在LUG蛋白中的功能具有重要的意义。
2.3 系统进化树分析
由于具有相同的氨基酸编码序列,所以我们仅选择了进行了系统进化树分析。根据系统进化树分析(图4),可将拟南芥和大白菜中的LUG蛋白分成Ⅰ和Ⅱ两个亚家族,每个亚家族都包含几个大白菜和拟南芥的基因,表明这一基因家族的基本特征在大白菜和拟南芥分离之前就已经形成,但是基因在这两组中的进化关系各不相同,如在I组中的和和II组中的和表现为旁系同源关系。在I组中的和、之间,以及Ⅱ组中的与、之间可以归类为直系同源进化基因。
3 讨论
本研究利用比较基因组学的方法,以拟南芥LUG蛋白家族基因为搜索对象,从大白菜基因组数据库中筛选到5个LUG蛋白家族基因。通过对拟南芥和大白菜LUG蛋白家族基因的序列保守性和系统进化分析发现,大白菜和拟南芥GIF蛋白家族之间存在较高的保守性,尤其在其N端Q-rich区和C端WD40重复区域更为明显,这种序列上的保守性提示大白菜LUG蛋白和拟南芥LUG蛋白之间可能具有功能上的相似性。
根据蛋白质序列进行了系统进化分析,每一亚族均包含拟南芥和大白菜LUG蛋白,这表明该基因亚族中的基因结构在拟南芥和大白菜分离之前就已经构建;同时在拟南芥和大白菜中还发现了一些旁系同源进化关系的基因,这表明LUG基因按照物种特异性的方式进行了一些扩张。
通过RNA可变剪接产生更多的蛋白种类,是生物体充分利用自身资源的一种有效的、经济的生物学方式[9-11]。拟南芥基因可能就是通过不同的RNA剪接产生。通过该方式,虽然产生了相同的蛋白编码序列(),但由于可变剪接所造成的5’ UTR区的差异同样会造成各成员间转录后调控上的差异,以达到更精细调控LUG蛋白质水平高低的目的,从而应对不同的生物学调节过程[12]。
本研究为今后阐明高等植物花器官发育过程中LUG蛋白的调控作用提供了一定的理论依据。随着对LUG蛋白研究的深入,LUG蛋白的功能将会被进一步了解,对LUG蛋白家族更加详尽的生物信息学分析以及利用分子生物学方法进一步阐明LUG蛋白在调控花器官发育和其它生物学程将是下一步的主要任务。


图4 拟南芥和大白菜LUG蛋白的系统进化树
[1] Coen E S, Meyerowitz E M. The war of the whorls: genetic interactions controlling flower development[J]. Nature, 1991, 353: 31-37
[2] Conner J, Liu Z. LEUNIG, a putative transcriptional corepressor that regulates AGAMOUS expression during flower development[J]. Proc Natl Acad Sci USA, 2000, 97: 12902-12907
[3] Liu Z, Meyerowitz E M.regulatesexpression inflowers[J]. Development, 1995, 121:975-991
[4] Drews G N, Bowman J L Meyerowitz E M. Negative regulation of thehomeotic geneby theproduct[J]. Cell, 1991, 65: 991-1002
[5] Roe J L, Nemhauser J L, Zambryski P C.participates in apical tissue formation during gynoecium development in[J]. Plant Cell, 1997, 9: 335-353
[6] Schneitz K, Hulskamp M, Kopczak S, et al. Dissection of sexual organ ontogenesis: a genetic analysis of ovule development in[J]. Development, 1997, 124: 1367-1376
[7] Liu Z, Franks R G, Klink V. Photoreceptors in signal transduction: pathways of enlightenment[J]. Plant Cell, 2000, 12: 1-14
[8] 杨红玉,张学琴. 拟南芥WD40蛋白[J]. 植物生理学通讯, 2008, 44(5): 1025-1033
[9] Black D L. Mechanisms of alternative pre-messenger RNA splicing[J]. Annu Rev Biochem, 2003, 72: 291-336
[10] Stamm S, Ben-Ari S, Rafalska I, et al. Function of alternative splicing[J]. Gene, 2005, 344: 1-20
[11] Lareau L F, Green R E, Bhatnagar R S, et al. The evolving roles of alternative splicing[J]. Curr Opin Struc Biol, 2004, 14: 273-282.
[12] Chung B, SimonsC, Firth A E, et al.Effect of 5'UTR introns on gene expression in[J]. BMC Genomics, 2006, 7: 120-132.
BIOINFORMATIC ANALYSIS OF LUG PROTEIN FAMILY INAND
MA Zong-qi1, WANG Feng-de2,*QIU Nian-wei1
(1.College of Life Science, Qufu Normal University, Qufu, Shandong 273165, China; 2.Vegetable Research Institute, Shandong Academy of Agricultural Sciences, Ji’nan, Shandong 250100, China)
LUG protein family is a kind of WD40 super family genes which features the existence of 7 repeat WD40 domains. The members of this family play an important role in the development of flower. In this study,bioinformatics analysis identified 7 and 5genes ins and Chinese cabbage, respectively.The conserved sequences of these proteins were analyzed and a polygenetic tree was constructed based on the corresponding cystatin proteins fromand Chinese cabbage. The results suggested that, besides the AtLUG6 gene contains 5 repeat, all the othergenes all contain 7 repeat WD40 domains. The polygenetic analysis of these sequences indicated that they could be divided into two sub-family groups and the main character forgenes might have existed before the split of Chinese cabbage and.
; Chinese cabbage; LUG protein; WD40 domain; bioinformatic analysis
Q344+.13
A
10.3969/j.issn.1674-8085.2012.04.010
1674-8085(2012)04-0042-04
2012-02-24;
2012-03-27
国家自然科学基金项目(31101553);山东省博士基金项目(BS2010SW027)
马宗琪(1964-),男,山东曲阜人,高级实验师,主要从事植物生理学教学与研究(E-mail: mazongqi640106@163.com);
王凤德(1979-),男,山东临沂人,博士后,主要从事蔬菜生理研究((E-mail:wfengde@163.com);
*邱念伟(1976-),男,山东嘉祥人,副教授,博士,硕士生导师,主要从事植物生理学教学与研究(E-mail:nianweiqiu@163.com).
