香溪河库湾春季水华纵向分布对水层结构的响应
2012-10-21刘德富黄钰铃王从锋
刘 流 刘德富 黄钰铃 王从锋
(三峡大学 水利与环境学院,湖北 宜昌 443002)
春季水华在富营养化水体的普遍暴发是一个世界性难题,各国学者已做了大量研究.Sverdrup[1]在1953年提出了解释海洋春季水华暴发的经典临界层理论,首次提出了水温分层诱发春季水华的概念模型,并成功应用于解释湖泊和水库中水华暴发和浮游植物群落结构演替过程[2-4].三峡水库支流库湾是典型的深水河道型库湾,对其春季水华暴发的特点和诱因,前人也已开展了大量研究.易仲强[5]发现香溪河库湾春夏期间存在显著的水温分层,徐耀阳[6]和王岚[7]发现香溪河库湾水华的暴发具有空间异质性,并根据纵向差异将香溪河库湾自下游往上游划分为干流区、湖泊区、过渡区和河流区[8].生态分区可以为支流水华的防控提供重要参考.王玲玲[9]得出库湾沿程浮游植物生物量与上游来流流量呈显著负相关,提出通过调节上游来流流量来控制支流水华.
然而,他们都忽略了香溪河库湾分层异重流的存在[10].分层异重流作为一个重要驱动力,极大地改变了库湾水体的理化分布格局[5,11],其在支流库湾的潜入深度、潜入厚度和影响范围将随水温分层的季节性变化和三峡水库的水位调度而改变,反过来导致支流库湾水温分层的纵向差异,从而直接或间接影响水华暴发情势[11].因此,不能对香溪河库湾进行简单的纵向分区.为验证以上理论,本研究于2010年春季水华期间对香溪河库湾的水动力、水温、营养盐等环境因子进行了现场监测,研究分层异重流对库湾水温分层的影响,并试图解释库湾春季水华浮游植物纵向分布差异的主导因素.
1 材料与方法
香溪河是三峡水库库首区域最大的一条支流,流域总面积3 095km2,长94km,多年平均流量为47.4 m3/s[12].它起源于位于湖北省西北部的神农架国家森林公园,向南流经兴山县和秭归县,在香溪镇注入长江.2003年6月三峡水库初次蓄水后,距香溪河与长江干流交汇点26km范围以内即被回水淹没,形成一个典型的深水河道型库湾.本研究于2010年4月18日对香溪河库湾进行了现场监测,三峡水库干流设置一对照点CJXX,库湾设点XX00~XX10,在未被库湾回水淹没的天然河道设置一对照点XXYT,如图1所示.
采用马骏[13]提出的三峡水库支流库湾低流速条件下的测流方法测香溪河库湾流速.获得的流速数据包括3个方向:东(Ve),北(Vn),垂向(Vu).水平向的流速由Ve和Vn投影在河流主泓线上合成,本研究忽略了垂向上的流速.用水质多参数仪(Hydrolab,美国HACH公司)测量水深、水温和电导率剖面.采用萨氏透明度板测量水体透明度,IL1400A水下光量子仪测量光合作用有效辐射(400~700nm).现场取350mL水样2瓶带回实验室参考《水和废水分析方法》(第4版)[14]测定水体总氮(TN)、总磷(TP)和溶解性硅酸盐(D-Si)及叶绿素a(Chl.a)浓度.

图1 香溪河库湾采样点分布图
真光层深度(Zeu)一般定义为水下光合作用有效辐射为水面的1%处对应的深度[15].为简化计算或弥补水下光强数据缺失,也可通过建立Zeu与水体透明度(SD)的关系式来计算Zeu.本文采用Zeu=1.802 1SD+1.818 2作为计算Zeu的公式[16].水体表面混合层深度(Zmix)是水体上层垂直混合充分的水层.本文采用水温垂向梯度不超过0.2℃/m时对应的深度计算.
2 结果分析
2.1 分层异重流特性
三峡水库干流水体的化学成分与支流库湾存在显著差别,电导率作为水体中溶解质浓度的表征,可用来示踪倒灌异重流的运动过程.图2给出了4月18日香溪河库湾的异重流沿纵剖面分布(白色箭头表示水流流入库湾,黑色箭头表示水流流出库湾;背景为电导率等值线,白色表示高电导率,黑色表示低电导率).整体来看,香溪河库湾的流速极低.三峡水库干流水体由表层30m处倒灌进入香溪河库湾,在距干支流交汇点约2km处表层异重流开始下潜为中层异重流,至距干支流交汇点约2km处异重流分为两部分,大部分水体以中层异重流形式继续下潜,小部分仍以表层异重流形式运行直至库湾回水末端.异重流两部分厚度均沿程减小.上游来流以底部异重流形式潜入库湾,在库湾中游约20km处出现分叉,一部分形成中层异重流,另一部分仍以底部异重流形式流出库湾.

图2 香溪河库湾异重流纵剖面分布
2.2 水温及光学分布特性
异重流水体一般混合比较均匀,因此异重流潜入深度的水温结构会发生改变.香溪河库湾水温垂向分布如图3所示.

图3 香溪河库湾水温垂向分布
由图3可以看出,三峡水库干流水体基本不存在水温分层(最大水温垂向梯度仅为0.07℃/m),而香溪河库湾下游XX00水温分层极其微弱,表层异重流水体水温几乎均匀分布(最大水温垂向梯度仅为0.09℃/m);XX01~XX02河段呈现相同的垂向水温结构,由于异重流在该河段自中层潜入,异重流的存在导致水体沿垂向在表层出现强制性分层(XX01处10 m以下最大水温垂向梯度仅为0.06℃/m);XX03自10m以下水温分层不显著(最大水温垂向梯度仅为0.14℃/m),10m以上的表层水温呈现较大梯度(最大水温垂向梯度为0.35℃/m),XX04出现同样的垂向水温结构,水温垂向梯度转折点出现在15m;库湾中游XX05~XX06河段受分层倒灌异重流影响,水温沿垂向出现双温跃层结构;XX07~XX08河段已不受中层倒灌异重流的影响,沿垂向呈现显著的水温分层(平均水温垂向梯度为0.27℃/m),但受表层倒灌异重流影响,表层5m以上呈现不同的水温梯度(5m以下平均水温垂向梯度仅为0.11℃/m),库湾上游XX09~XX10河段呈现同样的水温结构.受分层异重流影响,香溪河库湾呈现独特的水温分层结构,缺乏一般湖库水温分层出现的典型温跃层.
海洋和湖泊水体混合层深度一般取决于水温分层及风的 季节性 变化[18-19].卢 晨晨[20]对三峡库区河陆风规律的研究发现,三峡库区地面平均风速小于2 m/s;且香溪河库湾地形狭窄,风的最大传播距离不超过2km.因此,风对香溪河库湾的Zmix影响不显著,香溪河库湾Zmix的变化主要取决于水温分层的发育.图4给出了2010年4月18日香溪河库湾的Zmix纵向分布.可以看出,由于水温分层不显著,三峡水库干流和干支流交汇点附近的Zmix很大,几乎到底,自XX01至库湾回水末端Zmix呈现近似V字型分布,库湾中游XX05处Zmix达到11.97m.这是由于原有的水体混合层深度与上游异重流潜入深度相接,加深了水体混合层.图4也给出了Zeu的纵向分布,Zeu范围为2.72~8.49m,自库湾下游往回水末端递减.
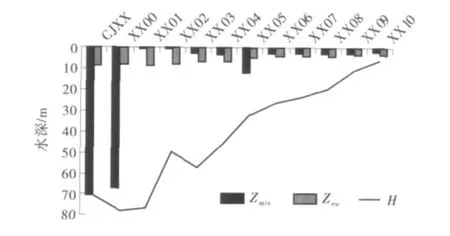
图4 香溪河库湾Zmix和Zeu纵向分布
2.3 营养盐分布特性
图5给出了2010年4月18日三峡水库干流和香溪河库湾的主要营养盐浓度纵向分布.可以看出,干支流TN浓度值整体较高,在0.68~1.90mg/L范围内波动;TN的纵向分布大致呈V字型,两端高、中上游XX07~XX09河段较低,最大值出现在XX10;而库湾上游来流TN浓度却低于XX10.TP整体值也偏高,在0.09~0.53mg/L范围内波动;TP大致自下游往上游呈递增趋势,回水末端XXYT最高.
氮磷比是衡量浮游植物生长是否受到氮或磷相对限制的一个重要指标,TN/TP=16时浮游植物生长不受氮或磷的相对限制[21].由图5可以看出,干流及库湾回水氮磷比均小于16,在5.21~13.56范围内波动,沿纵向大致呈递减态势,在上游回水末端略有上升.D-Si浓度在三峡水库干流较高,而在香溪河库湾中下游XX00~XX04河段均呈现较高浓度(大于7.00mg/L),自XX05至库湾回水末端整体呈U字型分布,D-Si浓度在0.16~5.10mg/L范围内波动,中上游XX06~XX08河段D-Si最低(均小于0.5 mg/L);库湾上游来流D-Si浓度(7.00mg/L)与中下游河段处于同一水平.
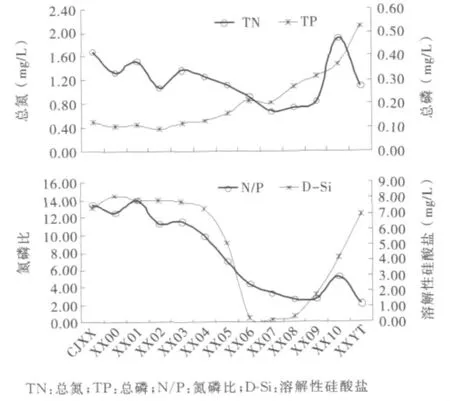
图5 三峡水库干流和香溪河库湾主要营养盐纵向分布
可以看出,三峡水库干支流的TN、TP浓度整体均较高,满足浮游植物生长需要,而其空间分布差异性较大.正如吉小盼[11]指出的,倒灌异重流对TN逆向补给效应显著.受昭君镇生活污水排放影响,回水末端XX10河段TN浓度显著高于中上游和上游来流.而由于香溪河库湾流域为富磷矿区,上游来流TP浓度极高(0.56mg/L),TP在库湾的纵向分布受上游来流影响显著.氮磷比的纵向分布显示,整个干支流均存在潜在的氮限制,而在库湾中上游尤为显著.上游来流与水库干流D-Si浓度基本持平,中上游河段低D-Si浓度表明硅藻水华繁殖消耗了大量溶解性硅酸盐.
3 讨 论
图6给出了干支流Zeu/Zmix和Chl.a的纵向分布,Zeu/Zmix是一个判别光照在垂向的分布和水体在垂向上的掺混对藻类生长的影响的综合指标[22],Chl.a常被用来表征浮游植物生物量[23].可以看出,Zeu/Zmix在水库干流及干支流交汇点附近均很小(0.12),下游河段 XX01~XX04呈现较大值,其中XX01、XX02均大于11;库湾中游XX05处出现Zeu/Zmix极小值(0.38),中上游河段XX06~XX10的Zeu/Zmix值在1.41~1.88范围内波动.Chl.a浓度的纵向分布存在显著差异,三峡水库干流Chl.a浓度极低,香溪河库湾中下游河段XX00~XX04浓度值范围为0.94~2.10mg/m3,而自 XX05至回水末端 Chl.a浓度沿程剧增,最大值高达161.41mg/m3,暴发了严重的水华.

图6 三峡水库干流和香溪河库湾
关于浮游植物生物量对水温分层的响应,已有学者做了大量研究.在营养盐、水温等因子满足浮游植物生长条件下,若水体出现较强温度分层,Zeu/Zmix>1时,浮游植物生长不受光照限制,这时候就会暴发高强度水华;反之,若Zeu/Zmix<1,即在水温分层不显著时,浮游植物颗粒就会被掺混至真光层以下,其接受光照的机会大大减小,生长就受到光限制[24].但若Zeu/Zmix≫1,即Zmix极小,营养盐受水温分层阻隔无法进入水体混合层,Zmix内的营养盐会很快消耗殆尽,无垂向迁移能力的浮游植物生长会受到营养盐限制[25];与之相反,Zeu/Zmix存在一个阈值,约处于0.2~0.35[24]之间(春季取0.35),小于这个阈值,由于浮游植物生物量的积累小于损失,则不可能暴发水华.因此,Zeu/Zmix=1是水华暴发的最有利条件[25].
香溪河库湾的营养盐条件对水华暴发无绝对限制,只在库湾中上游存在潜在的氮、硅限制,然而中上游却暴发了高强度的水华,显然无法用王岚[8-9]对支流库湾的分区理论来解释.而库湾Zeu/Zmix的纵向分布差异可以很好的解释这一现象,三峡水库干流和干支流交汇点附近Zeu/Zmix<0.35,因此Chl.a浓度极低;而XX01~XX04河段Zeu/Zmix≫1,由图7可以看出,该河段以硅藻、绿藻等无浮力或中性浮力藻类为主,这些藻类在高强度水温分层(Zmix很小)条件下,极易发生沉降损失,因此Chl.a浓度也极低;库湾中游XX05点Zeu/Zmix>0.35,之后中上游XX06~XX10河段Zeu/Zmix>1,暴发了高强度的硅藻水华.图7显示,甲藻只在香溪河库湾干支流交汇点附近XX00和回水末端XX10出现,这与甲藻喜高掺混水流密切相关;绿藻在三峡水库干流至库湾中游均存在,而硅藻主要出现在XX04~XX10,并在XX06~XX10河段占优.这也解释了图5中库湾中上游D-Si浓度偏低的现象.

图7 三峡水库干流和香溪河库湾浮游植物群落结构纵向分布
而由图4可知,Zeu/Zmix值主要由Zmix决定,因此,香溪河库湾水华情势纵向分布的差异主要由水温分层的差异决定,也在很大程度上受分层异重流的潜入深度和运行距离影响.而本文限于篇幅,只研究了香溪河库湾的一次春季水华,因此,未来有必要对香溪河库湾的分层异重流运动规律和浮游植物分布及演替过程进行长系列观测,探讨不同水库运行期及水库调度过程下浮游植物纵向分布及群落演替规律,以建立水库调度与支流库湾水华纵向分布的响应关系,为三峡水库的生态调度提供支撑.
4 结 论
1)春季香溪河库湾出现显著的分层异重流,水库干流水体以表中层异重流形式倒灌进入库湾,而上游来流首先以底部异重流形式潜入库湾,随后形成中层异重流.
2)分层异重流极大地改变了香溪河库湾的水温结构,水库干流水温不分层,而支流香溪河库湾中受复杂的分层异重流影响出现独特的水温垂向分层结构,缺乏典型的温跃层,且在中游出现双温跃层形式.
3)三峡水库干流和香溪河库湾水华暴发均不存在营养盐绝对限制,水库干流通过倒灌异重流对库湾的TN进行补给,而库湾上游来流是库湾高TP浓度的重要原因,库湾中上游受氮相对限制较显著.
4)香溪河库湾春季以硅藻水华为主,受水温分层影响,香溪河库湾Zeu/Zmix值沿纵向分布呈现极大差异性,从而导致浮游植物生物量和群落结构的纵向分布差异,而分层异重流导致这种差异的根本驱动力,因此,弄清异重流运行规律及特性对库湾水华的研究至关重要.
[1] Sverdrup H.On Conditions for the Vernal Blooming of Phytoplankton[J].ICES Journal of Marine Science,1953,18(3):287-295.
[2] Ganf G,Oliver R.Vertical Separation of Light and A-vailable Nutrients as a Factor Causing Replacement of Green Algae by Blue-Green Algae in the Plankton of a Stratified Lake[J].The Journal of Ecology,1982,70(3):829-844.
[3] Berger S A,Diehl S,Stibor H,et al.Water Temperature and Mixing Depth Affect timing and Magnitude of Events During Spring Succession of the Plankton[J].Oecologia,2007,150(4):643-654.
[4] Lindenschmidt K E,Chorus I.The Effect of Water Column Mixing on Phytoplankton Succession,Diversity and Similarity[J].Journal of Plankton Research,1998,20(10):1927-1951.
[5] 易仲强,刘德富,杨正健,等.三峡水库香溪河库湾水温结构及其对春季水华的影响[J].水生态学杂志,2009,2(5):6-11.
[6] Xu Y,Cai Q,Ye L,et al.Asynchrony of Spring Phytoplankton Response to Temperature Driver Within a Spatial Heterogeneity Bay of Three Gorges Reservoir,China[J].Limnologica-Ecology and Management of Inland Waters,2011,41(3):174-180.
[7] Wang L,Cai Q,Tan L,et al.Longitudinal Differences of Phytoplankton Community during a Period of Small Water Level Fluctuations in a Subtropical Reservoir Bay(Xiangxi Bay,Three Gorges Reservoir,China)[J].International Review of Hydrobiology,2011,96(4):381-396.
[8] Wang L,Cai Q,Zhang M,et al.Longitudinal Patterns of Phytoplankton Distribution in a Tributary Bay under Reservoir Operation[J]. Quaternary International,2011,244(2):280-288.
[9] 王玲玲,戴会超,蔡庆华.香溪河生态调度方案的数值模拟[J].华中科技大学学报:自然科学版,2009,37(4):111-114.
[10]纪道斌,刘德富,杨正健,等.三峡水库香溪河库湾水动力特性分析[J].中国科学:G辑,2010,40(1):101-112.
[11]吉小盼,刘德富,黄钰铃,等.三峡水库泄水期香溪河库湾营养盐动态及干流逆向影响[J].环境工程学报,2010,4(12):2687-2693.
[12]黄真理,李玉梁.三峡水库水质预测和环境容量计算[M].北京:中国水利水电出版社,2006.
[13]马骏,刘德富,纪道斌,等.三峡水库支流库湾低流速条件下测流方法探讨及应用[J].长江科学院院报,2011,28(6):30-34.
[14]王心芳,魏复盛,齐文启.水和废水监测分析方法[M].4版.北京:中国环境科学出版社,2002.
[15]Kirk J T O.Light and Photosynthesis in Aquatic Eco-systems[M].Cambridge University Press,1994.
[16]Liu L,Liu D F,Johnson D M,et al.Effects of Vertical Mixing on Phytoplankton Blooms in Xiangxi Bay of Three Gorges Reservoir:Implications for Management[J].Water Research,2012,46(7):2121-2130.
[17]Lawrence I,Bormans M,Oliver R,et al.Physical and Nutrient Factors Controlling Algal Succession and Biomass in Burrinjuck Reservoir[R].Land & Water Resources Australia,2000.
[18]Kling G W.Comparative Transparency,Depth of Mixing,and Stability of Stratification in Lakes of Cameroon,West Africa[J].Limnology and Oceanography,1988,27-40.
[19]Farmer D M,Carmack E.Wind Mixing and Restratification in a Lake Near the Temperature of Maximum Density[J].Journal of Physical Oceanography,1981,11(11):1516-1533.
[20]鲁晨晨.三峡坝区河陆风的理论与观测研究[D].南京:南京信息工程大学,2009:5.
[21]Goldman J C,Mccarthy J J,Peavey D G.Growth Rate Influence on the Chemical Composition of Phytoplankton in Oceanic Waters[J].1979,279:210-215.
[22]Jensen J,Jeppesen E,Olrik K,et al.Impact of Nutrients and Physical Factors on the Shift from Cyanobacte-rial to Chlorophyte Dominance in Shallow Danish Lakes[J].Canadian Journal of Fisheries and Aquatic Sciences,1994,51(8):1692-1699.
[23]Boyer J N,Kelble C R,Ortner P B,et al.Phytoplankton Bloom Status:Chlorophyll a Biomass as an Indicator of Water Quality Condition in the Southern Estuaries of Florida,USA[J].Ecological Indicators,2009,9(6):S56-S67.
[24]Oliver R,Hart B,Olley J,et al.The Darling River:Algal Growth and the Cycling and Sources of Nutrients[R].Murray Darling Basin Commission,Project M386 1999,2000.
[25]Reynolds C.Succession and Vertical Distribution of Phytoplankton in Response to Thermal Stratification in a Lowland Mere,with Special Reference to Nutrient A-vailability[J].The Journal of Ecology,1976,64(2):529-551.
[26]Lawrence I,Bormans M,Oliver R,et al.Factors Controlling Algal Growth and Composition in Reservoirs:Report of Reservoir Management Workshops January 2000[R].LWRRDC,Canberra,2000.
