基于CLIMEX和ArcGIS的槟栉盾蚧在中国的潜在地理分布研究
2012-09-28李志红黄冠胜吴杏霞吕文诚
方 焱, 李志红*, 黄冠胜, 吴杏霞, 傅 辽, 吕文诚
(1.中国农业大学农学与生物技术学院,北京 100193;2.国家质量监督检验检疫总局,北京 100088)
槟栉盾蚧 [Hemiberlesia rapax (Comstock)],又名桂花栉圆盾蚧[1],隶属于同翅目(Homoptera),盾蚧科(Diaspididae),栉圆盾蚧属(Hemiberlesia),英文名为greedy scale。该虫目前在我国尚无分布[2],是我国进境水果检疫所关注的有害生物之一[1],如2007年《中华人民共和国国家质量监督检验检疫总局和南非共和国农业部关于南非输华葡萄植物检疫要求议定书》将槟栉盾蚧列为中方关注的有害生物,该虫也出现在我国台湾水果输大陆检疫性有害生物名单上[3]。
该虫的寄主较为广泛,主要为害猕猴桃、山核桃、柑橘等重要水果,而且随植物产品进入中国的可能性较高,一旦传入将对我国的水果产业带来严重损失,也会影响我国进口贸易。因此,明确槟栉盾蚧在我国的潜在地理分布以及未来气候变化对其在中国潜在地理分布的影响,对科学制定检疫措施以防御其入侵具有重要意义。
目前,常用的适生性研究工具主要有CLIMEX、GARP、Maxent、DIVA-GIS 4种,本研究采用CLIMEX模型对槟栉盾蚧的潜在地理分布进行预测。至今,CLIMEX模型已被众多学者所采用,如墨西哥按实蝇在我国的适生性分析[4],番石榴果实蝇在我国的潜在地理分布[5],基于CLIMEX的橘实锤腹实蝇在中国的适生性分析[6],蜜柑大实蝇在中国的适生性[7]等研究均使用了CLIMEX工具。
本研究根据槟栉盾蚧的已知地理分布及相关生物学特性,运用适生性分析软件CLIMEX和地理信息系统ArcGIS相结合的方法,对该虫在我国目前及未来的潜在地理分布进行了预测。
1 材料与方法
1.1 基础资料
1.1.1 已知地理分布
通过对槟栉盾蚧相关文献资料查阅整理,结合CABI(Crop Protection Compendium,2011)[1]上的相关信息,得到槟栉盾蚧在全球的分布信息,槟栉盾蚧在美洲(百慕大群岛、墨西哥、美国、巴巴多斯、哥斯达黎加、古巴、危地马拉、洪都拉斯、牙买加、波多黎各、特立尼达和多巴哥、阿根廷、玻利维亚、巴西、智利、哥伦比亚、厄瓜多尔、圭亚那、秘鲁、乌拉圭),大洋洲(澳大利亚、新喀里多尼亚、新西兰、巴布亚新几内亚),欧洲(意大利、葡萄牙、俄国、西班牙、英国),非洲(南非、阿尔及利亚、肯尼亚、马达加斯加、马拉维、葡萄牙、塞舌尔、坦桑尼亚、赞比亚、津巴布韦),亚洲(印度、伊朗、伊拉克、日本、马来西亚、巴基斯坦、斯里兰卡)五大洲均有分布,目前在我大陆尚无发生报道。
1.1.2 生物学特性
槟栉盾蚧常在寄主树干的树皮处和寄主的枝条处发生,也常在树叶和果实上发生。在新西兰,新西兰琼楠属植物是槟栉盾蚧最主要的替代寄主[8]。
本研究根据槟栉盾蚧相关文献的查找、整理和分析,发现槟栉盾蚧主要在热带和亚热带地区发生。槟栉盾蚧的发育起点温度为9.3℃[9],其生长繁殖的最适温度在16.7~24.6℃之间,在温度为28℃时,槟栉盾蚧就不能产卵,它分别需要257、552、882日·度和992日·度才能进入第2、第3龄期,成熟态和生殖阶段[10],槟栉盾蚧生长最适湿度为75%[11]。
1.1.3 寄主与危害
寄主植物:槟栉盾蚧主要为害葡萄(Vitis vinifera)、猕猴桃属(Actinidia)、柑橘属(Citrus)等重要水果。它的寄主较为广泛,不仅包含水果果实,还包括景天属植物(Sedum)、梨树(Pyrus)、白杨(Populus)[1,12]等植物。
危害:槟栉盾蚧是一种为害植物果实和木质两部分的有害生物,主要在热带和亚热带地区发生。被槟栉盾蚧感染的植物会引起叶片变黄,叶片提前脱落,顶梢枯死等严重症状[8]。
1.1.4 传播途径
槟栉盾蚧主要借助风、鸟、或者其他介体昆虫进行传播[8]。
1.1.5 气候数据
本研究在2006年发布的CLIMEX 3.0版本中,导入了由Darren J.Kriticos提供的TYN SC 2.0数据集,该数据集包含了1961年到1990年的30年每月平均降雨量、每月平均温度、每日温度范围以及蒸汽压等关键气候数据,并且在这些变量的基础上,推算出了1961年到1990年间每月日最高温平均值、每月日最低温平均值、每月上午9点到下午3点的相对湿度平均值等气候数据。
秦大河于《中国气候与环境演变》一书对未来全球气候变化做出了预测[13],预测结果是与1961年到1990年30年的温度与降雨量的平均值相比较而得出的。2020年,中国气候的年平均气温将上升1.3~2.1℃,年平均降雨量将增加2%~3%;2050年,中国气候的年平均气温将增加2.3~3.3℃,年平均降雨量将增加5%~7%[13],本研究所采用的是上述预测结果的平均值。
1.1.6 地理数据
根据CABI(2011)官方网站发布的相关信息,并结合国内外相关文献,明确了槟栉盾蚧在全球的地理分布情况。地图资料包含中国国界及省界图(比例尺为1∶4 000 000),从中国基础地理信息系统网站(http:∥nfgis.nsdi.gov.cn)[14]下载得到。
1.2 分析工具及方法
本研究使用了中国农业大学植物检疫与入侵生物学实验室购买的CLIMEX 3.0和 ArcGIS 9.3正版软件,以CLIMEX和ArcGIS相结合的方法进行槟栉盾蚧在中国的潜在地理分布预测。因为槟栉盾蚧寄主广泛,本研究中不考虑寄主的影响。
2 结果
2.1 槟栉盾蚧适生性评判标准的确定
通常CLIMEX认为0<EI≤10时,物种低度适生;EI>20时,表示物种高度适生,EI介于两者之间则为中度适生[15-17]。我国学者使用该标准分析了番石榴果实蝇[5]、橘实锤腹实蝇[6]、纳塔尔实蝇[18]、枣实 蝇[19]、扶 桑 绵 粉 蚧[20]等 在 我 国 的 适 生性。本研究根据以上标准的同时,综合了槟栉盾蚧的实际地理分布情况。槟栉盾蚧在美国东部沿海的分布较为广泛,发生较为严重,本研究结果中,东部沿海的宾夕法尼亚州,弗吉尼亚州,北卡罗来纳州,南卡罗来纳州的EI值均大于20,因此本研究亦采用以上分级标准。
2.2 槟栉盾蚧适生性参数的确定
根据CLIMEX自带模板所提供的参数值及槟栉盾蚧的生物学资料,初步确定了槟栉盾蚧CLIMEX分析所需的各项参数,然后根据槟栉盾蚧在北美洲的实际分布区(图1),反复调试参数使槟栉盾蚧的预测分布范围(图2)最大程度的与实际分布相吻合,并以此参数对槟栉盾蚧在欧洲的分布区进行预测,预测结果较好地包含该地区的所有已知分布点,最终确定了槟栉盾蚧适生性分析的17个CLIMEX参数(如表1)。
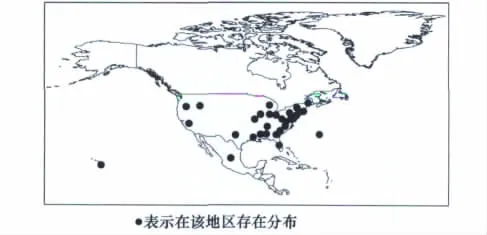
图1 槟栉盾蚧在北美洲的实际分布区图
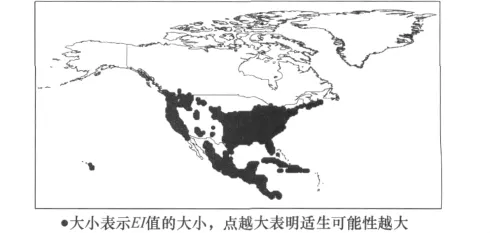
图2 基于CLMEX预测的槟栉盾蚧在北美洲的分布图
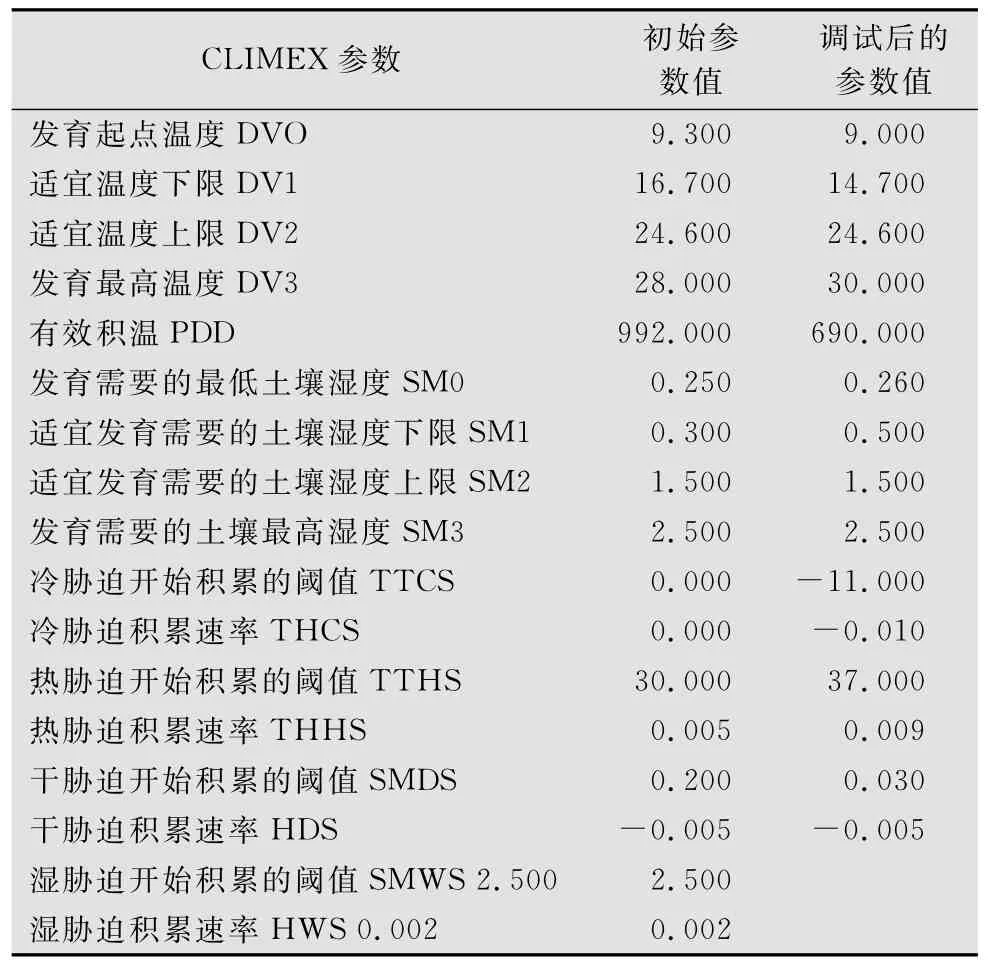
表1 槟栉盾蚧的地点比较模型参数
2.3 目前气候条件下槟栉盾蚧在中国的潜在地理分布
从图3中可以看出:海南、台湾、浙江、福建、广东、广西、贵州、重庆、湖南、湖北、江西全境,江苏、安徽、陕西、河南、四川、云南大部分地区,以及辽宁、山东、山西、宁夏、甘肃、西藏少部分地区为高度适生区,EI值均在20以上;山东大部分地区以及河南、河北、山西、陕西部分地区,甘肃、宁夏、四川、西藏零星地区为中度适生区,EI值在10~20之间;北京、天津全境,辽宁、河北大部分地区以及山西、陕西、宁夏、甘肃、四川、西藏少部分地区,云南零星地区为低度适生区,EI值在0~10之间;其他地区为槟栉盾蚧非适生区,EI值为0。
2.4 未来气候条件下的槟栉盾蚧在中国的潜在地理分布
从槟栉盾蚧2020年在中国潜在适生分布图(图4)和2050年在中国潜在适生分布图(图5)可以看出:2020年和2050年槟栉盾蚧在中国的适生区范围比当前气候条件下的适生范围都有所增大。
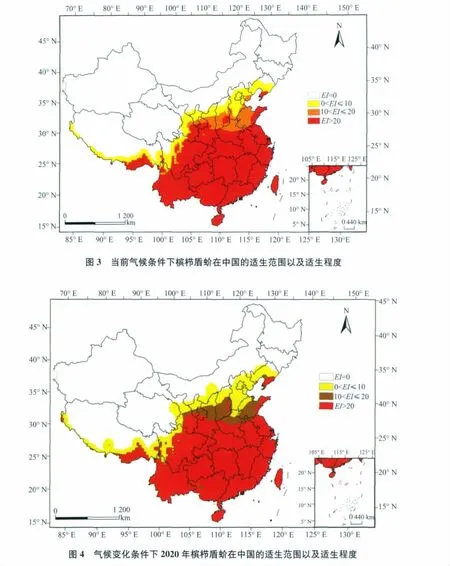
在2020年气候条件下,适生范围向北扩展。其中,浙江、福建、广东、贵州、重庆、湖南、江西、湖北全境,江苏、安徽、河南、陕西、四川、海南、台湾、云南大部分地区,以及山东、辽宁、西藏、甘肃少部分地区,山西、宁夏零星地区为高度适生区,EI值均在20以上;山东、山西大部分地区,河南、陕西、宁夏、甘肃少部分地区以及辽宁、四川、西藏、广西、台湾、海南零星地区为中度适生区,EI值在10~20之间;北京、天津全境,辽宁、河北、宁夏大部分地区,山东、河南、内蒙古、山西、陕西、甘肃、青海、四川、西藏少部分地区为低度适生区。2050年适生范围继续向北扩展,适生范围继续增大,吉林、新疆、内蒙古少部分非适生区变为低度适生区。
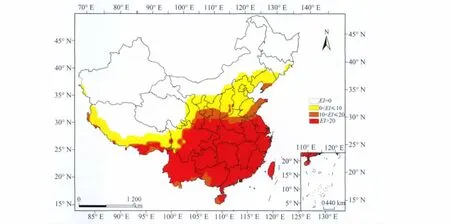
图5 气候变化条件下2050年槟栉盾蚧在中国的适生范围以及适生程度
2.5 目前和未来气候条件下槟栉盾蚧在中国的适生范围比较
如图6示,在目前气候条件下,槟栉盾蚧在我国的适生区面积达42.48%,其中低度适生区占6.67%,中度适生区占4.09%,高度适生区占31.72%;至2020年,随着气候的变化,适生区范围向北移动了一定的距离。槟栉盾蚧在我国的适生区面积增加至46.49%,其中,低度适生区占9.23%,中度适生区占4.56%,高度适生区占31.70%。2050年,适生区面积增至49.1%,其中低度适生区占14.56%,中度适生区占5.43%,高度适生区占29.11%。总体趋势是适生性面积增大,其中低度适生区和中度适生区都有所增大,但是高度适生区反而有一定程度的减少,高度适生区适生性降低。
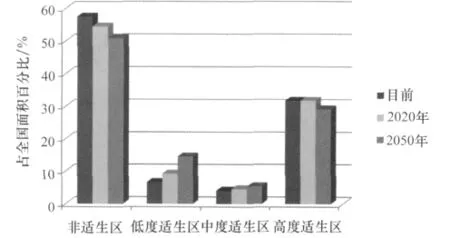
图6 目前及未来气候条件下槟栉盾蚧在中国的适生面积比较
具体的变化可以从图3~5中看出来,主要表现在:
(1)高度适生区:2020年与目前相比,辽宁、西藏、甘肃、四川、云南高度适生区有所增加。海南、台湾、广西、西藏零星高度适生区变为中度适生区,山东、山西、陕西部分高度适生区变为中度适生区。整体上高度适生区面积减少,在2020年变化的基础上,2050年高度适生区面积继续减少。
(2)中度适生区:2020年与目前相比,辽宁、山西、陕西、宁夏,甘肃中度适生区有所增加,其中山西、海南、台湾中度适生区主要是由高度适生区变为中度适生区而增加的,其他基本为低度适生区变为中度适生区。山东、河北、河南中度适生区范围有所减少,中度适生区变为低度适生区。整体上看,中度适生区面积增大,2050年,中度适生区面积进一步增大。
(3)低度适生区:2020年与目前相比,山东、河南、河北、内蒙古、青海低度适生区有所增加,其中,山东、河南、河北为中度适生区变为低度适生区导致低度适生区增加,内蒙古和青海低度适生区的增加是由于非适生区变为低度适生区。整体上看,低度适生区面积增大,2050年,低度适生区面积进一步增大。
总体上,在未来气候变化条件下,槟栉盾蚧在中国的适生范围有所增加,北方部分非适生区变为适生区,南方部分地区的适生性有所下降,适生范围向北扩展,我国更加适合槟栉盾蚧的生存。
3 讨论
3.1 槟栉盾蚧的生物学信息
关于槟栉盾蚧的生态学要求,目前的研究尚不深入。通过文献查找,发现关于槟栉盾蚧的研究内容主要包括部分分布信息、温度、积温的研究[1,9-11,21-23],以 及 寄 主 植 物 对 其 抗 性 研 究[11]、分 子水平进化学的研究[24]等,温湿度、积温等参数的研究尚不完全。缺乏文献数据的支持,增添了CLIMEX参数调试的难度。希望在今后的研究中,相关研究者对槟栉盾蚧进行更多的基础生物学研究,从而提高其适生性分析的可靠性。
3.2 未来气候对槟栉盾蚧潜在地理分布影响研究
温度上升是未来全球气候变化的主要特征之一,气候变暖会使有害生物发生危害的格局发生变化,危害区扩大[25]。受气候变暖的影响,昆虫倾向于向高纬度(两极方向)或高海拔分布扩散[26]。中国幅员辽阔,中国南部及东南部沿海地区生物入侵发生最为严重,气温限制了许多入侵种向北的扩散,而西北、华北和东北地区的持续变暖,将可能使某些重大入侵物种扩散至北方地区[27]。夏季高温也是限制昆虫分布的因素之一。当温度接近耐受范围上限,昆虫在低海拔、温暖地区的分布会适当收缩[28]。本研究中槟栉盾蚧的适生范围向北扩展,适生范围增大正符合以上论断。同时,南方部分地区的高度适生区的适生性有所下降,也正好验证了夏季高温限制了昆虫分布的论断。因此本研究对未来气候条件下槟栉盾蚧在中国的适生区预测结果较为合理。
3.3 管理措施建议
研究结果表明,国内存在槟栉盾蚧的寄主较为广泛,而且该虫在中国较易定殖,一旦传入我国,将对我国的水果产业带来消极影响。所以,要对该虫实施严格的检疫措施,防止该虫入侵我国。本研究认为应加强下述三方面的工作:(1)加强针对槟栉盾蚧的检疫措施。建议在相关水果的进口贸易中,加强原产地种植期间的监测与控制,在进口口岸严格把关,做好进口水果的检疫许可,现场检验、实验室检测、除害处理以及检疫监管等相关程序。(2)加强针对槟栉盾蚧的疫情监测。建议挑选浙江、福建、广东、江苏、陕西、山东等高度适生区的水果主产区,定点设立疫情监测点,一旦发现视情况加大监测范围,定期上报监测结果,形成针对槟栉盾蚧疫情的实时监控体系。(3)加强针对槟栉盾蚧的检疫宣传和教育。建议通过公共媒体,开展相关的检疫宣传,加大公共教育力度,进一步提高公众对槟栉盾蚧及其潜在危害的认识,做到全民防疫。
[1] 梁广勤.中国进出境水果关注的有害生物[M].北京:中国农业出版社,2009:28-29.
[2] CABI.Crop protection compendium[EB/OL].(2011-09-26)http:∥www.cabi.org/cpc/.
[3] 李今中,张晓燕,陈艳,等.台湾进境水果有害生物风险分析研究[J].中国农学通报,2009,25(22):228-235.
[4] 耿建,李志红,万方浩,等.墨西哥按实蝇在我国的适生性分析[J].植物保护,2008,34(4):93-98.
[5] 吕文刚,邓裕亮,李志红,等.番石榴果实蝇在我国的潜在地理分布[J].植物保护学报,2010,37(6):529-534.
[6] 倪文龙,陈洪俊,曲伟伟,等.基于CLIMEX的橘实锤腹实蝇在中国的适生性分析[J].植物检疫,2010,24(4):20-25.
[7] 王俊伟,李志红,陈洪俊,等.蜜柑大实蝇在中国的适生性研究[J].植物检疫,2009,23(1):1-4.
[8] Blank R H,Olson M H,Bell D S.Invasion of greedy scale crawlers(Hemiberlesia rapax)onto kiwifruit from taraire trees[J].New Zealand Entomologist,1987,10(1):127-130.
[9] Blank R H,Gill G S C,Upsdell M P.Greedy scale,Hemiberlesia rapax (Hemiptera:Diaspididae),phenology on kiwifruit leaves and wood[J].New Zealand Journal of Crop and Horticultural Science,1996,24(3):239-248.
[10] Blank R H,Gill G S C,Kelly J M.Development and mortality of greedy scale(Homoptera:Diaspididae)at constant temperatures[J].Environmental Entomology,2000,29(5):934-942.
[11] Hill M G,Mauchline N A,Connolly P G,et al.Measuring resistance to armoured scale insects in kiwifruit (Actinidia)germplasm[J].New Zealand Journal of Crop and Horticultural Science,2010,38(2):69-85.
[12]Schuh J,Mote D C.Insect pest of nursery and ornamental trees and shrubs in Oregon[R].Agricultural Experiment Station Bulletin,1948,449:1-164.
[13] 秦大河,丁一汇,苏纪兰,等.中国气候与环境演变·上卷[M].北京:科学出版社,2005:9-10.
[14] 中国基础地理信息系统[EB/OL].http:∥nfgis.nsdi.gov.cn.
[15] Robert W S,Gunter M A.climate model of the red imported fire ant,Solenopsis invicta Buren (Hymenoptera:Formicidae):Implications for invasion of new regions,Particularly O-ceania[J].Environmental Entomology,2005,34(2):317-335.
[16] Michael S W,Darren J K,Samantha A,et al.The hosts and potential geographic range of Dothistroma needle blight[J].Forest Ecology and Management,2009,257(6):1505-1519.
[17] Sutherst R W,Maywald G F,Bottomley W,et al.CLIMEX Version 3:User’s guide[EB/OL].http:∥www.Hearne.com.au,2009.
[18] 张哲,万方浩,李志红,等.利用CLIMEX预测纳塔尔实蝇在中国的潜在地理分布[J].生物安全学报,2011,20(1):43-49.
[19] 吕文刚,林伟,李志红,等.枣实蝇在中国适生性初步研究[J].植物检疫,2008,22(6):343-347.
[20] 马俊,胡学难,彭正强,等.基于CLIMEX模型的扶桑绵粉蚧在中国潜在地理分布预测[J].植物检疫,2011,25(1):5-8.
[21] Charles J G,Hill M G,Allan D J.Releases and recoveries of Chilocorus spp.(Coleoptera:Coccinellidae)and Hemisarcoptes spp.(Acari:Hemisarcoptidae)in kiwifruit orchards:1987-93[J].New Zealand Journal of Zoology,1995,22(3):319-324.
[22] Greaves A J,Davys J W,Dow B W,et al.Seasonal temperatures and the phenology of greedy scale populations (Homoptera:Diaspididae)on kiwifruit vines in New Zealand[J].New Zealand Journal of Crop and Horticultural Science,1994,22(1):7-16.
[23] Hill M G,Mauchline N A,Stannard K A.Predicting armoured scale insect(Homoptera:Diaspididae)phenology on kiwifruit(Actinidia sp.)[J].New Zealand Journal of Crop and Horticultural Science,2008,36(4):253-262.
[24] Geoffrey E M,Benjamin B N.A molecular phylogenetic study of armoured scale insects(Hemiptera:Diaspididae)[J].Systematic Entomology,2006,31:338-349.
[25] 戈峰.应对全球气候变化的昆虫学研究[J].应用昆虫学报,2011,48(5):1117-1122.
[26] 董兆克,戈峰.温度升高对昆虫发生发展的影响[J].应用昆虫学报,2011,48(5):1141-1148.
[27] 吴刚,戈峰,万方浩,等.入侵昆虫对全球气候变化的响应[J].应用昆虫学报,2011,48(5):1170-1176.
[28] Ponti L,Cossu Q A,Gutierrez A P.Climate warming effects on the Olea europaea-Bactrocera oleae system in Mediterranean islands:Sardinia as an example[J].Global Change Biology,2009,15(12):2874-2884.
