敲低Aurora A基因增强替莫唑胺抗人脑胶质瘤U87细胞的作用☆
2012-09-17陈云鹏张弩丁之明夏之柏
陈云鹏 张弩 丁之明 夏之柏
正常细胞分裂过程的任何偏差会造成中心体扩增、染色体异常分离以及非整倍体的产生,导致基因组不稳定,从而诱发肿瘤[1]。近年来,一种新的参与有丝分裂的丝氨酸苏氨酸激酶家族——Aurora家族逐渐被认识,其分为3类:Aurora-A、Aurora-B 和 Aurora-C,其中 Aurora-A 最受关注[2]。研究发现Aurora A过表达不但使某些肿瘤如卵巢癌的侵袭性和转移率明显提高,而且与患者预后直接相关[3-4]。敲低 Aurora A 在喉癌细胞的表达可使其对顺铂的敏感性增加[5]。我们先前的研究也发现Aurora A在胶质瘤中存在过表达现象。在此基础上,本文研究敲低Aurora A基因联合替莫唑胺(temozolomide,TMZ)治疗后细胞增殖变化,探讨其作为肿瘤基因治疗新靶点的可能性。
1 材料与方法
1.1 研究对象 U87细胞购自中国科学院细胞研究所;DMEM培养基、胎牛血清、胰蛋白酶购自美国Gibco公司;抗Aurora A抗体购自美国Bethyl公司;抗β-actin抗体、辣根过氧化物酶耦联的抗兔IgG和抗鼠IgG购自武汉博士德;替莫唑胺由美国Schering-Plough公司提供;二甲基亚砜(dimethyl sulfoxide,DMSO) 购自美国 Sigma 公司;pshRNA Lentivector质粒、包装质粒、基于人免疫缺陷病毒骨架的慢病毒包装系统 (Lenti-Pac HIV Expression Packaging Kit)购自美国GeneCopoeia公司;中量质粒DNA提取试剂盒购自德国Qiagen公司;人胆囊收缩素八肽(CCK8)购自美国Invitrogen公司。
1.2 细胞培养 U87细胞培养于含10%新生牛血清的高糖DMEM培养液,内含青霉素、链霉素,置37℃、5%CO2条件下常规培养。
1.3 细胞转染 构建Aurora A shRNA质粒载体(广州复能基因公司协助构建)。按说明书用Lenti-Pac HIV Expression Packaging Kit慢病毒包装系统进行慢病毒感染。慢病毒感染24 h后对样品进行换液处理;48 h后用荧光显微镜检查GFP表达;96 h开始添加浓度为7.5 μg/L的嘌呤霉素进行筛选。荧光显微镜下观察转染了Aurora A shRNA和空载体的U87细胞是否有荧光。细胞分未转染组(U87)、 空载体组 (U87-GFP) 及转染组(U87-shRNA)
1.4 RT-PCR检测Aurora A mRNA表达 提取3组细胞总RNA,按Promega试剂盒说明进行反转录和PCR扩增。Aurora A上游5′-AATCTGGAGGCAAGGTTCGACTG-3′,下游 5′-TGGGAGATAAGT GGTTAAGGAGG-3′,GAPDH为内参照。产物经1.5%琼脂糖凝胶电泳,紫外线下检测并拍照。
1.5 Western blot检测 Aurora A蛋白表达 提取3组细胞总蛋白,测定浓度;蛋白样品(每泳道50 μg)行 7.5%SDS PAGE 电泳,100 mV 转膜。牛奶封闭,加入 Aurora A 抗体(1∶200)和 β-actin 抗体(1∶500)4℃过夜,次日加入 HRP 标记二抗(1∶5000),37℃1 h,洗膜后ECL发光,X胶片曝光、显影。
1.6 CCK8法检测细胞活性 替莫唑胺用DMSO配制成浓度为 100 mmol/L的溶液,-20℃保存,临用时用全培养液稀释至所需浓度。取对数生长期的未转染组、空载体组和转染组细胞接种于96孔培养板,细胞密度 2 × 104/mL,置 37℃、5%CO2条件下培养24 h,分别加入浓度依次为10、25、50、100、200 μmol/L 的 TMZ,每孔加 100 μL 并设置空白对照组及阴性对照组(二甲基亚砜),每个药物浓度设5个平行孔,继续培养72 h。另取未转染组、空载体组和转染组细胞,加入 50 μmol/L TMZ分别作用24、48、72 h。完成处理后每孔加入CCK8试剂10 μL,37℃继续培养 2 h,全自动酶标仪检测波长450 nm的吸光度OD值。细胞生长抑制率(%)=(1-加药组OD平均值/对照组OD平均值)×100%。实验重复3次。做对数曲线算出IC50值。
1.7 流式细胞仪分析细胞 取对数生长期的未转染组、空载体组和转染组细胞,用TMZ同前处理。处理完成后,将细胞收集于流式管中,PBS洗涤,70%冰乙醇固定,再用PBS洗涤2次,每管加入预先配制好的PI染液l mL,室温避光5 h,上机检测细胞周期。
2 结果
2.1 Aurora A转染U87细胞的鉴定 荧光显微镜下转染组和空载体组细胞带绿色荧光蛋白,表明转染成功(图1)。用ImageJ软件分析条带灰度。RT-PCR结果灰度值分别为:未转染组 (31023±926)、 空载体组 (30124±1074)、 转染组 (896±172),各组间的差异有统计学意义(F=2159.63,P<0.001),两两比较显示转染组灰度值低于其他两组(P < 0.001);Western blot结果灰度值分别为:未转染组 (39556±2306)、 空载体组 (39348±2738)、转染组(574±96),各组间的差异有统计学意义(F=589.36,P < 0.001),两两比较显示转染组灰度值低于其他两组(P<0.001)。实验结果表明相对于空载体组及未转染组,转染组的Aurora A mRNA和蛋白表达均降低(图2、3)。
2.2 细胞生长抑制的比较 表1显示阴性对照组(二甲基亚砜)内,未转染组、空载体组、转染组间无统计学差异(F=0.35,P > 0.05)。表1、图 4A 表明在一定TMZ浓度范围内各组细胞生长抑制率随着浓度的升高而增加,呈现浓度依赖性。经不同浓度的替莫唑胺作用不同时间后,转染组细胞的TMZ剂量反应曲线左移。经直线回归方程计算,未转染组、空载体组、转染组的半数有效抑制浓度(IC50) 分别为:(43.38±2.15)μmol/L、(43.76±1.52)μmol/L、(22.20±2.34)μmol/L,各组间差异具有统计学意义(F=184.06,P < 0.001),两两比较显示转染组半数有效抑制浓度(IC50)低于其他两组(P < 0.001)。用 50 μmol/L TMZ 处理各组细胞,相同时间点各组的差异均有统计学意义,两两比较显示,相同时间点下转染组的细胞抑制均高于空载体组和未转染组(P < 0.01)(表2、图 4B)。

图1 转染组和空载体组细胞(×200,荧光显微镜)。A:转染组(明场);B:转染组(荧光);C:空载体组(明场);D:空载体组(荧光)

图2 转染前后U87细胞Aurora A mRNA表达(RT-PCR)
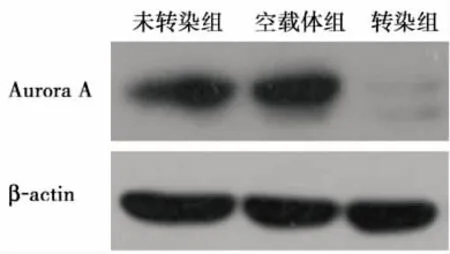
图3 转染前后U87细胞Aurora A蛋白表达(Western blot)

图4 替莫唑胺对未转染组、空载体组与转染组细胞增殖的影响
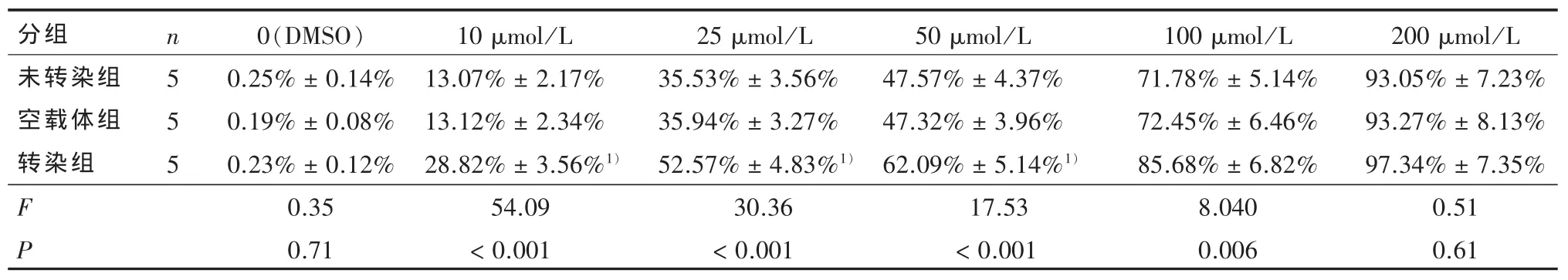
表1 不同浓度替莫唑胺作用于未转染组、空载体组与转染组细胞72 h的生长抑制率
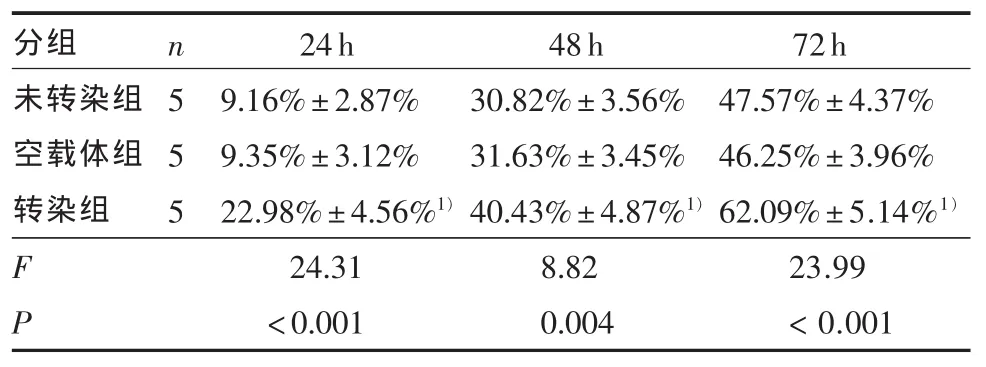
表2 未转染组、空载体组与转染组细胞在50 μmol/L替莫唑胺作用下不同时间点的细胞生长抑制率
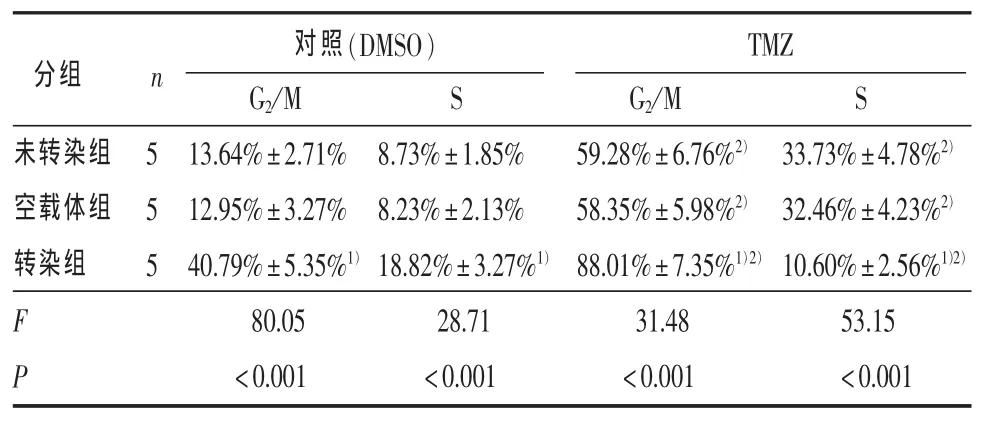
表3 未转染组、空载体组与转染组接受50 μmol/L替莫唑胺及对照(DMSO)处理72 h细胞周期变化(%)
2.3 细胞凋亡的比较 表3显示两种不同处理下未转染组、空载体组和转染组各组间G2/M期比例的差异均有统计学意义,两两比较显示两种不同处理下转染组G2/M期比例均高于其他两组 (P<0.01)。敲低Aurora A联合TMZ比单独敲低Aurora A或者单独 TMZ 处理 G2/M 期阻滞明显(P < 0.01)。
3 讨论
Aurora A位于重要的癌基因区域20q13,在有丝分裂期定位于中心体和纺锤体,在中心体复制、分离、成熟的过程中发挥重要作用。Aurora A高表达通过干扰纺锤体组装的检测点,引起胞质分裂中止,最终出现中心体扩增形成非整倍体细胞。少数非整倍体细胞生存下来,其生长优势高于邻近的正常细胞,导致形成肿瘤的可能性大大增加[6]。
我们发现Aurora A基因在胶质瘤细胞高表达,敲低其表达后,细胞增殖明显降低。Cammareri等[7]报道,直肠癌高表达Aurora A基因,沉默其表达后细胞增殖也明显下降,对5-FU的敏感性也明显增加。Lehman NL等[8]发现Aurora A过表达可促进喉鳞状上皮癌细胞的增殖和转移,敲低Aurora A基因表达可使喉鳞状上皮癌对顺铂的敏感性增加。我们发现敲低Aurora A基因不但使替莫唑胺对胶质瘤细胞的IC50明显降低,而且也可使细胞经较低浓度替莫唑胺作用后增殖活性显著下降。综上所述,敲低Aurora A基因表达不但能使多种肿瘤增殖活性下降,而且能增加肿瘤对化疗的敏感性。
Aurora A基因表达下调增加肿瘤细胞化疗敏感性的机理尚不清楚。我们的研究证实敲低Aurora A可使肿瘤细胞的G2/M比例增加,协同TMZ处理肿瘤细胞G2/M比例进一步增加,但凋亡并无增加。Marumoto等[9]在非霍奇金淋巴瘤中发现 Aurora A过表达破坏细胞G2/M检查点的功能,导致细胞周期无法阻滞于G2/M期;并使细胞在DNA受损情况下继续进行分裂,产生恶性转化。故敲低Aurora A的表达,可能是通过恢复细胞G2/M检查点的功能,使其得以发挥自身的监控作用,产生G2/M期阻滞,导致细胞生长抑制。Lee S 等[10]认为 Aurora A可对p53的315位丝氨酸磷酸化,导致其通过泛素化途径降解。敲低Aurora A的表达可能恢复p53功能,产生G2/M期阻滞,达到抑制增殖的作用。p53功能降低或缺失,bcl-2表达水平过高,以及表皮生长因子受体表达过高可干扰细胞对DNA损伤的反应,降低对化疗药物的敏感性[11]。诱导恶性胶质瘤细胞发生自噬是替莫唑胺的重要作用。由于自噬的特点,药物诱导肿瘤细胞产生自噬会出现两种不同的结果:一是保护肿瘤细胞免受放化疗损伤,另一种是启动细胞主动性的Ⅱ型细胞死亡程序。自噬早期抑制剂抑制替莫唑胺诱导自噬的同时可削弱其抗肿瘤效应,而自噬晚期抑制剂可诱导凋亡增加替莫唑胺抗肿瘤效应[12]。
Zhang J等[13]发现在替莫唑胺耐药细胞株中U373VR细胞中MGMT蛋白上调;SNB19VR细胞中的hMSH6表达缺失造成的MMR损伤。体外和临床试验研究均证实肿瘤的烷化剂抵抗性是由MGMT过表达或MMR缺陷介导的。而Aurora A的上调都可以使MGMT过表达或MMR缺陷介导,使肿瘤获得对替莫唑胺导致的损伤耐受或修复的能力。因此敲低Aurora A除可延长细胞有丝分裂G2期外还有下调MGMT修复MMR的功能,从而增强替莫唑胺的抗肿瘤作用。Aurora A的敲低不仅其本身就具有抗肿瘤作用,而且能增强肿瘤对替莫唑胺等化疗药物的敏感性,因此Aurora A基因将是未来胶质瘤基因治疗和联合化疗的重要靶基因。
[1]Endersby R,Baker SJ.PTEN signaling in brain: neuropathology and tumorigenesis[J].Oncogene,2008,27(41):5416-5430.
[2]Barton VN,Foreman NK,Donson AM,et al.Aurora kinase A as a rational target for therapy in glioblastoma[J].J Neurosurg Pediatr,2010,6(1):98-105.
[3]Yang SB,Zhou XB,Zhu HX,et al.Amplification and overexpression of Aurora-A in esophageal squamous cell carcinoma[J].Oncol Rep,2007,17(5):1083-1088.
[4]Landen CN Jr,Lin YG,Immaneni A,et al.Overexpression of the centrosomal protein Aurora-A kinase is associated with poor prognosis in epithelial ovarian cancer patients[J].Clin Cancer Res,2007,13(14):4098-4104.
[5]Loh JK,Lieu AS,Chou CH,et al.Differential expression of centrosomal proteins at different stages of human glioma[J].BMC Cancer,2010,10(7):268-295.
[6]Lehman NL,O′Donnell JP,Whiteley LJ,et al.Aurora A is differentially expressed in gliomas,is associated with patient survival in glioblastoma and is a potential chemotherapeutic target in gliomas[J].Cell Cycle,2012,11(3):489-502.
[7]Cammareri P,Scopelliti A,Todaro M,et al.Aurora-a is essential for the tumorigenic capacity and chemoresistance of colorectal cancer stem cells[J].Cancer Res,2010,70 (11):4655-4665.
[8]Lehman NL,O'Donnell JP,Whiteley LJ,et al.Aurora A is differentially expressed in gliomas,is associated with patient survival in glioblastoma and is a potential chemotherapeutic target in gliomas[J].Cell Cycle,2012,11(3):489-502.
[9]Marumoto T,Hirota T,Morisaki T,et al.A novel Aurora kinase A inhibitor MK-8745 predicts TPX2 as a therapeutic biomarker in non-Hodgkin lymphoma cell lines[J].Leuk Lymphoma,2012,53(3):462-471.
[10]Lee S,Cimica V,Ramachandra N,et al.Aurora A Is a Repressed Effector Target of the Chromatin Remodeling Protein INI1 /hSNF5 Required for Rhabdoid Tumor Cell Survival[J].Cancer Res,2011,71(9):3225-3235.
[11]张俊平,魏大年,史泓浏,等.体外药敏试验及化疗耐药和敏感分子特征指导的恶性脑胶质瘤临床个体化化疗研究[J].中国神经精神疾病杂志,2009,35(8):476-480.
[12]Lin CJ,Lee CC,Shih YL,et al.Resveratrol enhances the therapeutic effect of temozolomide against malignant glioma in vitro and in vivo by inhibiting autophagy[J].Free Radic Biol Med,2012,52(2):377-391.
[13]Zhang J,Stevens MF,Laughton CA,et al.Acquired resistance to temozolomide in glioma cell lines: molecular mechanisms and potential translational applications[J].Oncology,2010,78(2):103-14.
