牛膝菊属两种外来入侵植物叶片的形态解剖结构比较研究
2012-09-15昌恩梓齐淑艳孔令群李晓春
昌恩梓,齐淑艳,2,孔令群,李晓春,申 柳
(1.沈阳大学生物与环境工程学院,辽宁 沈阳 110044;2.辽宁省城市有害生物治理与生态安全重点实验室,辽宁 沈阳 110044)
牛膝菊属(Galinsoga)植物全世界约有5种,主要分布于美洲.我国有牛膝菊(Galinsoga parviflora)和粗毛牛膝菊(Galinsogaquadriradiata)2种,均为外来入侵植物,目前这2种植物在辽宁省都有分布[1-2].牛膝菊和粗毛牛膝菊作为一种入侵性杂草,其分布范围较广,生境条件多样,目前还不很清楚它们为什么具有如此强的入侵能力.植物体内部结构和外部特征是植物最为主要的性状,入侵植物对生境的适应和散布能力是以内部结构作为基础的,探讨入侵植物结构与生态适应性、入侵机理的关系,对入侵植物的防范和治理具有一定的意义.有关牛膝菊和粗毛牛膝菊的生理、生态学方面的研究在国内外已有报道[3-11],但对其叶的比较解剖学研究尚缺乏系统的材料.本文对分布于辽宁省的牛膝菊和粗毛牛膝菊叶片形态结构进行了较为系统的研究,以为防控入侵植物及入侵机理的研究提供科学依据.
1 材料与方法
1.1 供试材料
供试材料采自沈阳大学校园(N41°46′,E123°26′),本地区是沈阳市人口密集的商业文化中心地带,属暖温带半湿润季风型大陆性气候,四季分明,雨热同季.年平均气温为7.4℃,1月平均气温为-12.6℃,7月平均气温为27.5℃;年平均降水量为755.4mm,多集中在6—8月,可占全年降水量的64.4%;无霜期150d.在植物区系上,沈阳属于华北植物分布区,但正处于长白、蒙古植物分布区的交汇地带[12],植物种类比较丰富,给外来种入侵创造了生态环境条件.
2011年8月15日至9月6日9:00~11:00,于沈阳大学校区采集分布于不同草坪正常生长开花的牛膝菊(G.parviflora)和粗毛牛膝菊(G.quadriradiata).
1.2 研究方法
1.2.1 叶片形态特征的观测
分别取牛膝菊和粗毛牛膝菊新鲜植株各20株,每株取顶端成熟叶片至下数第3节完整、成熟的叶片2枚.利用清华紫光B700扫描仪将叶片形状扫描成图片文件,编号保存.将编号后的叶片分别装袋,80℃(24h)烘干至恒重,用电子天平(1/10000)称量每个叶片的干重.用AUTOCAD软件测量每片叶的长度、宽度、周长、面积;计算叶片的长宽比、叶形指数(叶面积/(长×宽))、比叶重(叶生物量/叶面积)[14].
1.2.2 气孔形态的观测
分别取牛膝菊和粗毛牛膝菊植株各30株,取其顶端成熟叶片至下数第3节完整、成熟的叶片,采用双刃刀片刮取叶片位于主脉两侧和近叶边缘的上、下表皮制作临时水装片.将制好的装片利用Motic(DMB52223IPL)数码显微镜摄像系统进行观测(10×40倍),每个装片随机选取3个视野进行拍摄,利用Motic mages Advanced 3.2图像处理软件,测量气孔(两个肾形保卫细胞的胞间隙)的长度、宽度、周长、面积及气孔数、表皮细胞数,计算气孔指数(100×每个视野单位面积内的气孔个数/(气孔个数+表皮细胞数))、气孔密度(个/mm2)、变异系数(标准差/平均数×100%)[4].
1.2.3 叶片解剖结构
将上述采集的新鲜叶片用刀片过中脉切成0.5cm×0.5cm左右的小片,FAA固定液固定,过夜.常规石蜡切片法制作叶的永久横切片[13],切片厚度10~12μm,酒精脱水,二甲苯透明,番红-固绿对染,加拿大树胶封片.每种植物叶片各制作有效石蜡切片30片.利用Motic(DMB52223IPL)数码显微镜摄像系统进行观测(10×10倍),每个装片随机选取3个视野进行拍摄,利用Motic mages Advanced 3.2图像处理软件,测量叶片表皮细胞厚度和宽度、栅栏组织长度和宽度、海绵组织厚度等,并计算栅栏组织与海绵组织的比值.
原始数据的整理采用Excel软件完成,差异显著性检测利用DPS软件进行.
2 结果与分析
2.1 叶片形态特征
牛膝菊和粗毛牛膝菊叶片形态特征测量结果见表1.分析结果表明,生长在相同的生境下,两种植物叶片的长宽比、叶形指数、单叶干重和比叶重差异不显著(P>0.05),而单叶面积表现为牛膝菊<粗毛牛膝菊,差异非常显著(P<0.01).

表1 牛膝菊和粗毛牛膝菊叶片的形态数量特征
2.2 叶表皮细胞形态特征
牛膝菊和粗毛牛膝菊的上、下表皮细胞均为不规则形,垂周壁呈波状,但上表皮呈浅波状,下表皮呈深波状.上、下表皮均有气孔分布,且下表皮多于上表皮,差异非常显著(P<0.01).保卫细胞为肾形,气孔器类型为不规则型(见图1).牛膝菊和粗毛牛膝菊叶片的气孔形态数量特征和气孔参数见表2.由表2可见,牛膝菊上表皮与下表皮气孔的长度、宽度和周长差异不显著(P>0.05);上表皮的气孔面积>下表皮的气孔面积,差异显著(P<0.05);而气孔指数和气孔密度均为上表皮<下表皮,差异非常显著(P<0.01).粗毛牛膝菊上表皮与下表皮气孔的长度、宽度和周长差异不显著(P>0.05);而上表皮的气孔面积<下表皮的气孔面积,差异显著(P<0.05);气孔指数和气孔密度均表现为上表皮<下表皮,差异非常显著(P<0.01).

图1 叶片表面观(10×40)

表2 牛膝菊和粗毛牛膝菊的气孔形态数量特征和气孔参数
2.3 叶片解剖结构
2.3.1 表皮
牛膝菊和粗毛牛膝菊的上、下表皮细胞均为一层,横切面呈长方形或椭圆形.粗毛牛膝菊表皮毛为3-细胞单列型(见图2).牛膝菊和粗毛牛膝菊的表皮细胞解剖特征见表3.数据分析表明:牛膝菊和粗毛牛膝菊上表皮细胞的厚度>下表皮的厚度,差异非常显著(P<0.01);上表皮细胞的宽度>下表皮的宽度,差异显著(P<0.05).
2.3.2 叶肉
牛膝菊和粗毛牛膝菊具有明显的栅栏组织和海绵组织分化,为异面叶,其解剖结构特征详见表3.栅栏组织细胞呈长柱状,1层位于近轴面(见图2).牛膝菊的栅栏组织细胞长度和宽度分别为72.20,23.82μm,海绵组织厚度为91.74μm,栅栏组织厚/海绵组织厚为0.7963,海绵组织细胞排列较疏松.粗毛牛膝菊栅栏组织细胞长度和宽度分别为72.85,29.85μm,海绵组织厚度为80.87μm,栅栏组织厚/海绵组织厚为0.9224,海绵组织细胞排列较紧密.
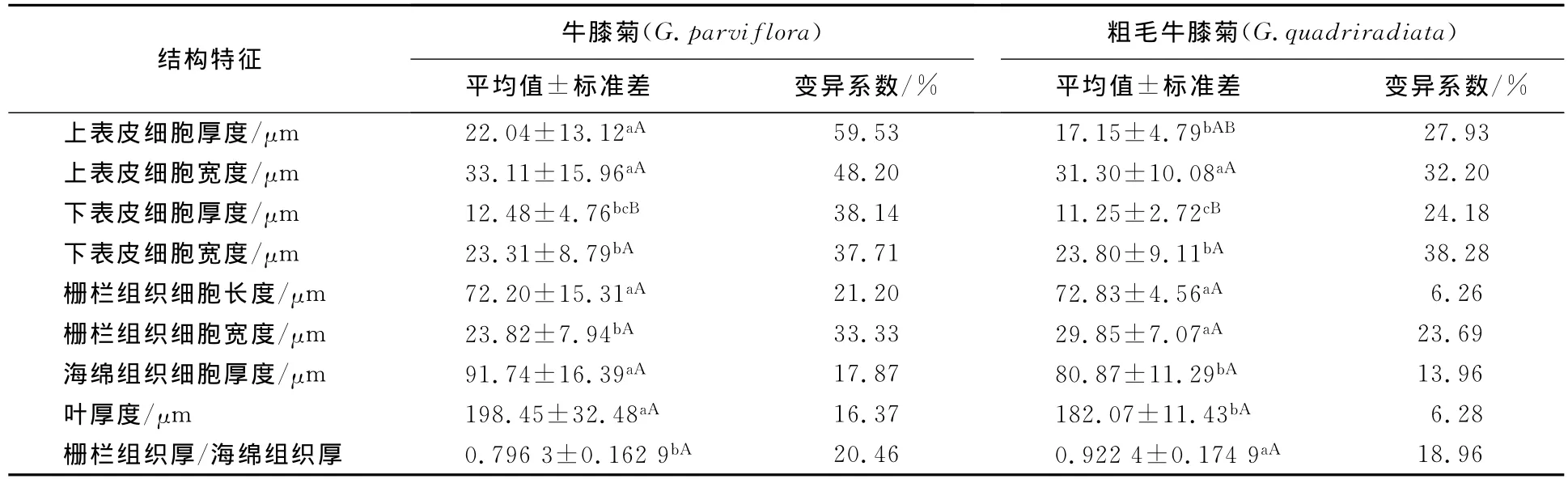
表3 牛膝菊和粗毛牛膝菊叶片的解剖特征
2.3.3 叶脉
牛膝菊和粗毛牛膝菊的中脉仅有1束维管束,外有2层维管束鞘,属于C3植物类型(见图2).
3 讨论
生态适应是生物个体为了适应环境变化,从形态结构、生理特征等方面做出的有利于生存的改变.叶片是植物体进行光合作用的营养器官,同时也是对环境变化最为敏感的器官,其形态结构特征最能体现环境因子的影响或植物对环境的适应[14].
3.1 叶片形态特征
入侵植物具有对异质性环境的快速适应能力和快速进化的特点[15].调查中发现,粗毛牛膝菊生境多样,而牛膝菊一般生长在蔽荫和水分充足的环境中.实验结果表明,粗毛牛膝菊的单叶面积(4.8890cm2)大于牛膝菊的单叶面积(2.9957cm2),大的叶面积具有增强光合作用效率、保持体内营养物质的作用,说明粗毛牛膝菊比牛膝菊能更好地适应资源贫瘠和干旱的环境,扩散能力更强,这也是目前粗毛牛膝菊分布多于牛膝菊的原因之一.而粗毛牛膝菊和牛膝菊植物叶片的长宽比、叶形指数差异不显著,表明生长在相同生境下同属不同种植物叶片的形态特征差别不大.
生物量和比叶重作为植物叶片的生长指标能够反映植物的生长和生产力大小.比叶重的增大可减少植物因叶表面蒸腾而引起的水分丧失,保水能力更强;此外比叶重增大可使叶片具有较强的支持和抵御能力[16].粗毛牛膝菊的比叶重(0.0030g/cm2)大于牛膝菊的比叶重(0.0003g/cm2),表明粗毛牛膝菊比牛膝菊具有更强的抗旱能力,这也是粗毛牛膝菊向西北和东北入侵潜力较强的原因之一.
3.2 叶表皮细胞形态特征
牛膝菊和粗毛牛膝菊上表皮气孔密度小于下表皮的,且差异非常显著,这样的气孔分布特点可使植物能够抵御强烈的日光照射、减少呼吸和蒸腾以达到降低水分散失的目的,说明牛膝菊和粗毛牛膝菊都是典型的阳生植物.野外观察表明,在沈阳霜降以后,其他植物地上部分均已死亡,而牛膝菊和粗毛牛膝菊仍能存活,表现出较强的抗寒能力.本实验的结果表明,牛膝菊和粗毛牛膝菊的上、下表皮细胞垂周壁呈波状,细胞更具弹性,可使其不受表皮细胞因寒冷失水所产生的压力影响[17],而植物叶片表皮细胞具有波浪状垂周壁,是抗寒能力较强的表现[18],这一结论与观察结果相吻合.
牛膝菊和粗毛牛膝菊气孔密度的变异系数都比气孔指数的变异系数大,说明在影响气孔参数因素相同的情况下,气孔指数是相对稳定的系统演替性状.这一结果与马清温等对北美红杉和水杉的气孔参数的研究结论一致[19-20].实验表明牛膝菊和粗毛牛膝菊的下表皮气孔密度和气孔指数的变异系数均大于上表皮,表明牛膝菊和粗毛牛膝菊叶片下表皮的气孔分布情况比上表皮更加稳定.但是粗毛牛膝菊的气孔面积小于牛膝菊的气孔面积,这种特点的气孔对外界条件变化反应灵敏,当植株受到干旱胁迫而叶片水分状况并未发生改变时,气孔导性下降,蒸腾降低,气孔可较快关闭,防止叶片可能发生的水分亏缺,从而提高植株的抗旱能力[21].在干旱环境下气孔密度一般随着环境中水分和湿度的减少而增加,但气孔面积则向小型化发展[22].粗毛牛膝菊气孔面积较小表现出对干旱环境的适应性要强于牛膝菊.
叶片表皮结构,特别是气孔参数,对环境条件变化的响应与适应多样、复杂,要深入揭示入侵植物叶表皮细胞形态特征对生态环境变化的适应性仍需要进一步开展相关的研究工作.
3.3 叶片解剖结构
牛膝菊和粗毛牛膝菊叶片上表皮细胞厚度均大于下表皮,栅栏组织与海绵组织的厚度比分别为0.7963和0.9224.栅栏组织与海绵组织的厚度比越大其耐寒、耐旱性越强,厚度比大于1的植物通常被认为是典型的抗寒、抗旱性植物[23].牛膝菊和粗毛牛膝菊栅栏组织与海绵组织的厚度比均较高,说明这两种植物的耐旱性、耐寒性都较强.而粗毛牛膝菊栅栏组织与海绵组织的厚度比大于牛膝菊的,且差异显著,表明在相同的环境条件下,粗毛牛膝菊比牛膝菊具有更强的光合能力,可以积累更多的有机物.粗毛牛膝菊的比叶重(0.0052g/cm2)大于牛膝菊的比叶重(0.0038g/cm2),且差异显著,也证明了这一点.海绵组织的厚薄主要反映了植物对水分条件的适应性,水分条件越好其海绵组织也就越厚[24],牛膝菊海绵组织厚度为91.74μm,粗毛牛膝菊海绵组织厚度为80.87μm,差异显著,说明粗毛牛膝菊对干旱环境条件的适应性强于牛膝菊.
以上叶片形态结构的特征反映出粗毛牛膝菊比牛膝菊适应范围更广、对逆境的忍耐力更强,能较好的适应干旱、寒冷环境,入侵潜力更大,这也是粗毛牛膝菊成功入侵的重要原因之一.同时这也说明水分和寒冷条件不能成为牛膝菊和粗毛牛膝菊等入侵植物继续扩大分布范围的限制因素.
[1]李振宇,解焱.中国外来入侵种[M].北京:中国林业出版社,2002:166.
[2]齐淑艳,徐文铎.外来入侵植物粗毛牛膝菊在辽宁地区新的发现[J].辽宁林业科技,2008(4):22-24.
[3]齐淑艳,徐文铎,文言.外来入侵植物牛膝菊种群构件生物量结构[J].应用生态学报,2006,17(12):2283-2286.
[4]齐淑艳,郭晓华,赵明.CO2、O3浓度升高对入侵植物牛膝菊叶片形态特征的影响[J].东北师大学报:自然科学版,2009,41(2):149-153.
[5]李康,郑宝江.外来入侵植物牛膝菊的入侵性研究[J].山西大同大学学报:自然科学版,2010,26(2):69-71.
[6]范雪涛,马丹炜,向莎,等.不同逆境条件下辣子草抗氧化酶系统的变化[J].应用与环境生物学报,2008,14(5):616-619.
[7]马丹炜,范雪涛,葛方兰,等.低温胁迫对辣子草水浸提液化感作用的影响[J].热带亚热带植物学报,2008,16(6):526-530.
[8]田陌,张峰.入侵物种粗毛牛膝菊(Galinsogaquadriradiata)在秦岭地区的生态适应性[J].陕西师范大学学报:自然科学版,2011,39(5):71-75.
[9]MERIC C,DANE F.A new alien species for the flora of European Turkey:Galinsogaparviflora (Asteraceae)[J].Phytologia Balcanica,2005,11:63-66.
[10]ESPINOSA-GARCIA F J,VAZQUEZ-BRAVO R,MARTINEZ-RAMOS M.Survival,germinability and fungal colonization of dimorphic achenes of the annual weed Galinsogaparviflora buried in the soil[J].Weed Research (Oxford),2003,43:269-275.
[11]FU DOSTATNY D.Effect of density on individual size and fertility in two species of weeds:Galinsoga ciliata and Galinsoga parviflora[J].Journal of Ecology,1999,47:353-357.
[12]徐文铎,何兴元,陈玮,等.沈阳市区植物区系与植被类型的研究[J].应用生态学报,2003,14(12):2095-2102.
[13]李正理.植物制片技术[M].北京:科学出版社,1987.
[14]王勋陵,王静.植物的形态结构与环境[M].兰州:兰州大学出版社,1989.
[15]BLOSSEY B,NOTZOLD R.Evolution of increased competitive ability in invasive non indigenous plants:a hypothesis[J].J Ecol,1995,83:887-889.
[16]HIGUCHI H,SAKURATANI T,UTAUNOMIYA N.Photosynthesis,leaf morphology and shoot growth as affected by temperatures in cherimoya(Annona cherimola Mill)trees[J].Sci Hort,1999,80:91-104.
[17]SANFORD W G.Pineapple croplay-concept and development[J].Batter Crop Plant,1962,46:32-43.
[18]陈清西,廖镜思,王明双,等.食用蕉若干品种类型叶片组织结构的比较观察[J].福建农林大学学报:自然科学版,1992,21(4):406-412.
[19]马清温,张金保.水杉(杉科)的叶表皮结构[J].植物研究,2003,23(1):32-35.
[20]马清温,李凤兰,李承森.气孔参数的变异系数和影响因素[J].北京林业大学学报,2005,27(1):19-23.
[21]李芳兰,包维楷,刘俊华.岷江上游干旱河谷海拔梯度上四川黄栌叶片特征及其与环境因子的关系[J].西北植物学报,2005,25(11):2277-2284.
[22]崔国文,马春平.紫花苜蓿叶片形态结构及其与抗寒性的关系[J].草地学报,2007,15(1):70-74.
[23]石登红,陈训.6种杜鹃花属(Rhododendron)植物叶片结构的研究[J].贵州科学,2005,23(3):39-45.
[24]容丽,陈训,汪小春.百里杜鹃属13种植物叶片解剖结构的生态适应性[J].安徽农业科学,2009,37(3):1084-1088.
