大叶藻(Zostera marina L.)胚的结构和种子萌发过程的研究*
2012-09-06田萍萍刘延岭崔翠菊梁广津钱冠兰王宏梅
田萍萍,刘延岭,崔翠菊,梁广津,李 霞,赛 珊,钱冠兰,王宏梅
(山东东方海洋科技股份有限公司,山东 烟台264003)
大叶藻(Zostera marina L.)胚的结构和种子萌发过程的研究*
田萍萍,刘延岭,崔翠菊,梁广津,李 霞,赛 珊,钱冠兰,王宏梅
(山东东方海洋科技股份有限公司,山东 烟台264003)
以采自山东荣成褚岛桑沟湾南岸成熟的大叶藻种子为材料,研究了种子的外部形态特征、胚乳成分及胚的解剖结构,并初步了解了大叶藻种子的萌发过程。结果表明,大叶藻种子是单子叶有胚乳的种子,呈暗褐色,椭圆形或长圆形,种皮革质且有17~20条纵肋,胚乳丰厚坚硬,胚乳细胞被淀粉粒充满;胚弯曲,由胚根、胚轴、胚芽和子叶组成,子叶U形棒状,其弯曲部有一裂缝,胚芽位于裂缝处,胚轴较短,其下是两个对生的胚根,有中央维管束贯穿胚根、胚轴、胚芽和子叶。在子叶出土萌发的植物类型中,大叶藻种子萌发过程是罕见的。萌发初期,子叶优先生长突破种皮而长出地面,此后胚根才开始生长并产生不定根。对大叶藻种子结构和萌发过程的研究,为探讨大叶藻种子萌发和幼苗更新的限制因素提供了一定的理论依据。
胚;萌发过程;海洋高等植物;种子结构;大叶藻
大叶藻(Zostera marina L.)是一年或多年生海洋高等植物,生于潮间带和潮下带的浅海中,营沉水生活。其形成的广阔海草场是近海生态系统重要的初级生产力,为许多生物提供重要的群落生境[1]。大叶藻是北半球沿海分布最广泛的海草,在东北亚、远东地区、欧洲与北美均有分布,国内主要分布于河北、山东、辽宁等省沿海[2-3]。近几十年,大叶藻草场在世界范围内大面积的消失直接导致了浅海生物栖息地的破坏,近海渔场的衰落及浅海水域生物多样性的丧失。这次世界范围的衰退在上世纪70-80年代起引发了全世界对这一生境的关注和研究。至今,在大叶藻研究领域内,修复生态学仍然是研究需求最为迫切最受关注的研究方向。
上世纪末,大叶藻草场的修复工作依赖于成株移栽。近年研究发现,用大叶藻种子播种较成株移栽更便于收集、储存与运输,大大降低了工作量及工作成本。因此,国内外学者开始将大叶藻草场的修复工作更多的寄望于种子。此时,对大叶藻种子基本生物学特性的深入了解,转为种子移栽研究开展的必要前提。
目前,有关大叶藻种子生物学特性的研究甚少,主要集中于种子萌发影响因子的研究[4-5],但萌发机制与环境因子的作用机理尚不清楚。对种子结构与成分的准确阐述,是了解种子适应环境条件生理过程的基础。原永党等[6]曾通过石蜡切片得到了大叶藻种皮棱角和胚乳的解剖结构。在此基础上,本研究首次利用石蜡切片得到大叶藻胚的解剖结构,并结合胚的解剖结构阐释了大叶藻种子的萌发过程。对大叶藻种子结构和萌发过程的研究,为探讨大叶藻种子萌发和幼苗更新的限制因素提供了一定的理论依据。
1 材料与方法
1.1 材 料
大叶藻种子于2009-08采自山东荣成褚岛桑沟湾南岸。采集后经过筛选,得到的成熟种子用自然海水冲洗干净,置于10℃海水中进行暂存。
1.2 方 法
1.2.1 大叶藻种子外部形态特征的观察
将大叶藻种子用尖头镊子固定在解剖镜载物台上进行外部形态特征观察。
1.2.2 大叶藻胚的石蜡切片
1.2.2.1 取材与固定
将用海水浸泡7~10d的成熟种子去掉种皮和胚乳,获得完整的胚。将胚放入FAA固定液固定48h。
1.2.2.2 梯度酒精脱水
固定好的胚依次入50%酒精、70%酒精、80%酒精、95%酒精和无水乙醇中做脱水处理,各级间隔2h。
1.2.2.3 透明、浸蜡、包埋
将脱水后的材料置入无水乙醇/二甲苯(1∶1)混合液30min,然后在二甲苯中进行透明处理约10min。
随后,把材料入二甲苯/石蜡(1∶1)混合液30min,再放入58℃熔融的纯蜡(Ⅰ)→纯蜡(Ⅱ)中各浸蜡45min。
将浸蜡后的材料放入恒温箱中保持58℃,用包埋剂进行常规石蜡包埋。
1.2.2.4 切片、展片
修整蜡块后,在RM2015型轮转式切片机上进行常规石蜡连续切片,切片厚度为10μm。将连续切片浮在40℃水浴锅中展片,用涂有甘油蛋白的载玻片附片后室温晾干。
1.2.2.5 脱蜡、复水
充分干燥的切片先后入二甲苯(Ⅰ)和二甲苯(Ⅱ)进行脱蜡处理20min和10min。
脱蜡后的切片依次入二甲苯/无水乙醇(1∶1)混合液、无水乙醇、95%酒精、80%酒精、70%酒精和50%酒精各5min进行复水。
1.2.2.6 苏木精-伊红染色
复水后的切片先用苏木精染色5min,然后依次入50%酒精、70%酒精和80%酒精各5min进行脱水处理。脱水后的切片依次入10%的伊红酒精溶液染色1min,然后依次入95%酒精(Ⅰ)各5min、95%酒精(Ⅱ)5min、无水乙醇(Ⅰ)5min,无水乙醇(Ⅱ)各5min洗去非特异性的着色。
1.2.2.7 封片、观察
将染色切片依次入二甲苯/无水乙醇(1∶1)混合液5min、二甲苯(Ⅰ)10min、二甲苯(Ⅱ)20min进行透明处理,并采用中性加拿大树胶封片,日产Olympus倒置显微镜观察并拍照。
1.2.3 胚乳的组织化学成分鉴定
将新鲜种子的胚乳做徒手切片,用I2-KI试剂滴染鉴定淀粉和蛋白质,呈蓝色为淀粉,呈黄色为蛋白质;用苏丹Ⅲ染色鉴定脂肪,呈红色为脂肪。
1.2.4 观察大叶藻种子萌发和幼苗形成的过程
为考察大叶藻胚各部分结构发育的历程,将部分成熟种子置于10℃无阳光直射的自然海水中,逐日观察大叶藻种子萌发和幼苗形成的过程。
2 结 果
2.1 种子的外部形态特征
大叶藻种子呈暗褐色,长约4mm,椭圆形或长圆形,合点端圆钝珠孔端有凹陷。种皮外层革质,上具清晰纵肋17~20条延伸至两端,内层膜质光滑易与胚乳剥离。
2.2 胚乳的成分鉴定
成熟的种子胚乳丰厚坚硬。由I2-KI试剂染色(图1),除少数胚乳细胞中空,其他胚乳细胞均被染成蓝色的淀粉粒充满,未见蛋白质和脂肪贮存。

图1 胚乳细胞经I2-KI试剂染色(×800)Fig.1 Endosperm cell stained with I2-KI reagent(×800)
2.3 胚的解剖结构
由大叶藻胚纵切片可见(图2a,2b),大叶藻胚弯曲,由胚根、胚轴、胚芽和子叶组成。子叶一片,U型棒状,占整个胚体体积和解剖结构的85%以上,而胚根、胚轴和胚芽位于子叶的弯曲部,仅占不足15%。由胚纵切图可见(图2c),胚根细胞与邻近的胚轴细胞表现为明显不同的外形。胚轴由较胚根细胞体积稍大的分生细胞构成,长度较短,两端分别连接胚芽与胚根,侧面连接子叶。
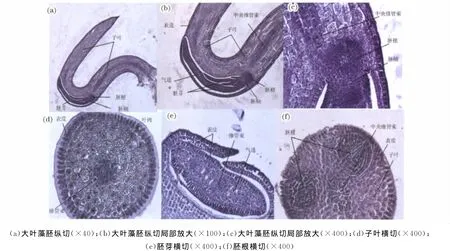
图2 大叶藻胚的石蜡切片Fig.2 Paraffin Section of Embryo of Zostera marina L.
由子叶横切可见(图2d),其外层表皮细胞排列致密,内部叶肉细胞体积大、核小、细胞质浓厚、胞间隙大,中央是由胚芽和胚根分生组织分化出的中央维管束。在胚纵切的连续石蜡切片中可见中央维管束贯穿胚根、胚轴、胚芽和子叶。
由胚芽连续横切切片可见(图2e),其最外层表皮细胞排列整齐,内部散布肉细胞、气道及中央有维管束。在胚芽的基部背向子叶的弯曲部一侧有一裂缝,是幼叶生长向外伸出的部位。
由胚根纵切可见(图2f),胚有两个对称排列的胚根,呈圆锥状,朝向表皮。其尖端生长点由体积小且排列紧密的分生细胞组成。细胞质浓厚、核大。生长点以上的细胞体积较大且伸长,整齐排列成伞状,列间隙较大。
2.4 大叶藻种子萌发和幼苗形成
在10℃无阳光直射的自然海水中,大叶藻种子经7~10d放置开始萌发。由图3可见,大叶藻种子的萌发从子叶开始,首先子叶伸长突破种皮,在伸长到可以接受足够光照的时候,胚芽发育而来成的2~3片嫩绿的幼叶才从子叶的裂缝处伸长并生长,其生长所需的营养物质来自胚乳。此时伸长的子叶仍是白色的,其与幼叶连接处有一胚轴发育而成的节点。此后胚根开始发育,在节点处长出一对对生根。种子携带的营养物质完全耗尽后,子叶与胚乳脱落,形成具有完整的根茎叶结构的幼苗。
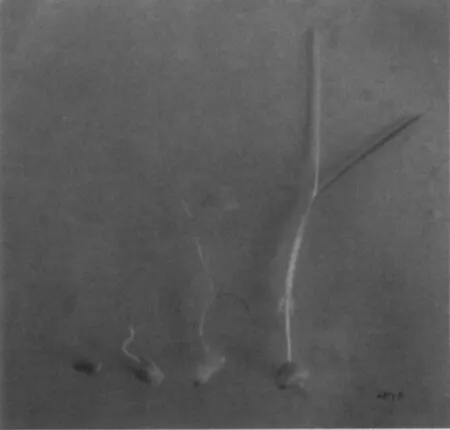
图3 大叶藻种子萌发过程ig.3 Seed germination of Zostera marina L.
3 讨 论
3.1 胚、胚乳和种皮对种子萌发的影响
种子在适宜条件下,是否能够萌发主要取决于种子的成熟度,
种子成熟应具备生理方面的成熟和胚形态的完全建成。成熟胚在形态上应分化形成完整的子叶、胚芽、胚轴和胚根四部分。实验观察大叶藻种子胚在形态上已分化成熟,在形态上不会影响种子的萌发。大叶藻种子是单子叶有胚乳的种子,胚乳中含有丰富的淀粉,为其胚的发育提供了足够的营养储备。
种子萌发有个吸水膨胀的过程,较坚硬的胚乳会导致种子的吸水过程缓慢,使萌发过程迟缓。大叶藻种子较坚硬的革质种皮会对种子萌发产生机械阻力,使种胚难以冲破种皮,发芽整齐度和出苗率降低。另外,革质的种皮会阻碍水分和气体的渗透,使胚得不到发芽所需的水分和氧气,而使种子强迫性休眠。所以,较坚硬的胚乳和革质的种皮可能是种子萌发的外部限制因素。
3.2 幼苗的形成
在子叶出土萌发的植物类型中,大叶藻种子萌发过程是罕见的。萌发初期,子叶优先生长,首先突破种皮长出地面,随后胚根才开始生长并产生不定根。Fishman和Orth[7],Orth等[8]曾报道大叶藻幼苗的成活率不足10%。自然条件下大叶藻营养体形成的冠层引起营养和光照的竞争,从而限制了幼苗生长,即使种皮可能经较长时间降解而萌发,幼苗成活率可能极低或不能成活。王孝民等[9]认为水深是限制多年生沉水植物微齿眼子菜(Potamogeton maackianus)幼苗成活的一个关键因素。随水深增加,光照减弱,幼苗真叶形成、不定根产生前,由于光合作用不足,导致营养缺乏而死亡。Van Wijk[10]认为沉水植物篦齿眼子菜(potamogeton pectinatus)的幼苗能长成成熟个体取决于幼苗所处地点的植被情况。本研究认为,大叶藻种子完全具有在潮间带或潮下带的浅海、植被稀疏的地点,产生幼苗进而长成成熟个体的较大潜力。在对幼苗生长的过程中观察发现,在子叶和胚乳还未脱落之前,围绕胚乳的外侧会长出一丛须状物,幼苗通过这种须状物交织在一起生长。在随海流扩散的过程中,遇到合适的基质可以附着后扎根生长,起固着器的作用,这也是与海洋沉水生活相适应的结果。
3.3 种子的休眠
种子是一个处于休眠状态的有生命的活体,它受内在或外在因素的限制,短时间内存在不能萌发或萌发困难的现象,这是植物对外界条件长期形成的一种适应性。大叶藻种子在不利环境下也具有休眠的特性,休眠时间可长达一年之久[8]。Van和 Verschuure[11]及 Meling-lopez和Ibarra-Obando等[12]曾报道在大叶藻的有性繁殖中,其种子产量在时间和空间上都是不可预知的。大叶藻属于海洋沉水种子植物,种子成熟后或落于海底沉积层上,或随水流冲到其它地方,在此过程中绝大部分被多种原因(菌类侵袭,动物捕食与破坏以及其他物理、化学条件的影响)破坏掉了,还有一部分处于休眠状态的种子则埋藏于海底沉积物中成为大叶藻种子库,在合适的干扰作用下对退化生态系统的恢复以及植被更新发挥重要的作用,对大叶藻种子休眠特性的研究,对种子保存和促萌发有很大的现实指导意义。
致谢:东方海洋科技股份有限公司国家海藻工程研究中心处对本研究进行过程中提供的帮助,李晓捷老师、罗世菊老师及江鑫博士等各位工作人员的指导和帮助。
(References):
[1] THAYER G W,BJORNDAL K A,OGDEN J C,et al.Role of larger herbivores in seagrass communities[J].Estuaries,1984,7(4A):351-376.
[2] YE C J,ZHAO K F.Advances in the study on the marine higher plant eelgrass(Zostera marina L.)and its adaptation to submerged life in sea water[J].Chinese Bulletin of Botany,2002,19(12):184-193.叶春江,赵可夫.高等植物大叶藻研究进展及其对海洋沉水生活的适应[J].植物学通报,2002,19(2):184-193.
[3] YU H,MA Y H,ZHANG Y,et al.Ecological characteristics of eelgrass(Zostera marina L.)and its response to environmental changes[J].Transactions of Oceanology and Limnology,2007,(suppl.):112-120.于函,马有会,张岩,等.大叶藻的生态学特征及其与环境的关系[J].海洋湖沼通报,2007,(增刊):112-120.
[4] MOORE K A,ORTH R J,NOWAK J F.Environmental regulation of seed germination in Zostera marina L.(eelgrass)in Chesapeake Bay:effects of light,oxygen and sediment burial[J].Aquatic Botany,1993,45(1):79-91.
[5] ABE M,KURASHIMA A,MAEGAWA M.Temperature requirements for seed germination and seedling growth of Zostera marinafrom central Japan[J].Fisheries Science,2008,74:589-593.
[6] YUAN Y D,SONG Z C,GUO C L,et al.Morphological characters and microstructure of Zostera marina[J].Transactions of Ocean-ology and Limnology,2010,3:73-78.原永党,宋宗诚,郭长禄,等.大叶藻形态特征与显微结构.海洋湖沼通报,2010,3:73-78.
[7] FISHMAN J R,ORTH R J.Effects of predation on Zostera marina L.seed abundance[J].Journal of Experimental Marine Biology and Ecology,1996,198:11-26.
[8] ORTH R J,HARWELL M C,BAILEY E M,et al.A review of issues in seagrass seed dormancy and germination:implications for conservation and restoration[J].Marine Ecology Progress Series,2000,200:277-288.
[9] WANG X M,SUN S G,GUO Y H.A primary study of germination characteristic of Potamogeton maackianus[J].Acta Hydrobio-logica Sinica,2003,27(1):95-97.王孝民,孙士国,郭友好.微齿眼子菜瘦果萌发特性的初步研究[J].水生生物学报,2003,27(1):95-97.
[10] WIJKV R J.Ecological studies on Potamogeton pectinatus L.Ⅲ.Reproductive Strategies and germination ecology[J].Aquatic Botany,1989,33(3-4):271-299.
[11] VAN L F,VERSCHUURE J M.Comparative study on populations of Zostera marina L.(eelgrass):experimental germination and growth[J].Journal of Experimental Marine Biology and Ecology,1995,185:77-91.
[12] MELING-LÓPEZ A E,IBARRA-OBANDO S E.Annual life cycles of two Zostera marina L.populations in the Gulf of California:contrasts in seasonality and reproductive effort[J].Aquatic Botany,1999,65:59-69.
(王佳实 编辑)
Study on Embryo Structure and Seed Germination of Zostera marina L.
TIAN Ping-ping,LIU Yan-ling,CUI Cui-ju,LIANG Guang-jin,LI Xia,SAI Shan,QIAN Guan-lan,WANG Hong-mei
(Shandong Oriental Ocean Sci-Tech Co.,Ltd,Yantai 264003,China)
Seed morphological character,endosperm components and embryo structure of the Zostera marina L.were studied using the materials collected from south bank of Sanggou Bay,Rongcheng Zhudao in Shandong province.The process of the seed germination was also studied in this paper.The results showed that Zostera marina L.is monocotyledon.The testa is leathern and has17-20longitudinal ribs.Its seed contains dark brown endosperm in ellipse or egg-shape.The endosperm is rich and hard,filled with starch grain.The embryo is curve and composed of radicle,hypocotyl,embryo bud and cotyledons.The cotyledons showed in U-shaped rod with embryo bud in its gap of the curved part.The hypocotyl is a little short and two opposite radicles were located underside.The central vascular bundle runs through the embryo.The seed germination process is exceptional among the epigaeous plants.In the early germination stage,the cotyledons break through the testa and grow up out of the ground,thereafter,the radicles begin to grow and produce adventitious roots.This study provided a theoretical basis for further studies on limiting factors of seed gemination and seedling of Zostera marina L..
embryo;germination;marine higher plants;seed structure;Zostera marina L.
August 25,2011
Q174
A
1671-6647(2012)03-0402-06
2011-08-25
国家海洋局开放基金——典型水域清洁生产与环境安全技术与示范(200905020-12);山东省科技发展计划项目——人工大
叶藻场构建的生物学研究(2010GSF10612);烟台市科学技术发展计划项目——人工大叶藻场构建的生物学研究(2009211)
田萍萍(1983-),女,山东德州人,硕士,工程师,主要从事海洋生物学方面研究.E-mail:pptian2010@163.com
