动植物分类任务中的左侧半球优势
2012-09-05王培培陈昭燃
王培培 陈昭燃
(首都医科大学高级脑功能研究中心,北京100069)
对外界信息的分类加工是人脑实现高效知觉加工的基本机制[1-2]。以此机制为基础,人脑发展出加工不同类别刺激的“专家化系统”,如对面孔信息具有特异性的梭状回面部区(fusiform face area,FFA)[3]。目前对自然物体分类的研究多采用动物/非动物或者人工物体作为刺激,也有许多研究[4-6]比较生物与非生物刺激。然而却很少有研究涉及“生物”这一大分类中的差异。生物进一步又可分为动物和植物,它们在人类进化和社会化过程中扮演的作用不同:动物对人类的威胁更具主动性和攻击性。在长期的进化中,人类大脑可能发展出对动植物不同的加工机制。例如人类视觉加工系统对运动物体具有明显的加工优势[7]。
在研究哪些脑区对动植物概念的加工存在差异前,一个重要的问题是:对动植物概念的加工是否存在半球优势?为验证这种可能性,我们分离动物类别和植物类别的刺激,测量被试判断它们的声音反应任务的反应时差异。进而,我们试图检验人类对动植物类别的加工机制是否与脑功能的偏侧化有关。我们比较右利手被试分别用左手和右手对动植物分类的按键反应。右利手被试通常存在左半球加工优势[8-9],如果人脑对动植物分类的加工存在偏侧化现象,那么对动植物判断在使用左手或右手做出反应时应存在差异。
1 材料与方法
1.1 研究对象
从首都医科大学招募25名右利手被试[女性,平均年龄(23.00±1.75)岁],所有被试视力或矫正视力正常,无神经、精神病史。被试在实验前阅读并签署了行为研究知情同意书。此研究获得首都医科大学伦理委员会的批准。
1.2 问卷测量
爱丁伯格利手量表[10]:测量被试在应对日常活动(如:写作、用筷子、刷牙、用剪刀)时使用右手或者左手的频率。被试根据实际情况评定自己在10个日常活动项目中的左右手习惯。评定规则为:若倾向于使用某一只手,在相应的框内打一个“+”;若某只手绝对占优,一般从来不试着使用另外一只手去尝试,在相应的框内打两个“+”;若无论是用哪只手都可以,在左右两栏中都打一个“+”。
特质心理测量量表[11-12]:包括特质焦虑量表(trait anxiety inventory,TA)和状态焦虑量表 (state anxiety inventory,SA)。TA描述相对稳定的,作为一种人格特质且具有个体差异的焦虑倾向;SA描述一种不愉快的情绪体验,如紧张、恐惧、忧虑和神经质,伴有非自主神经系统的亢进,一般为短暂性的。本实验使用这两种量表测量被试是否存在焦虑特质或处于焦虑状态。
1.3 刺激材料
研究者自《现代汉语频率词典》[13]选取120个单字名词,包括60个动物名词(如:猫、猪、狼)及60个植物名词(如:柳、莲、梅),并严格控制词频和笔画数,使其在两种类别间差异无统计学意义[t(59)=0.59,P=0.126]。这些词被制作为正方形黑底白字图片,水平和垂直视角均为1°,用于呈现给被试。
1.4 实验程序
被试坐在计算机屏幕(19英寸CRT显示器)正前方,距离屏幕80cm。实验前要求被试填写上述量表,并练习实验任务。在正式实验的每个trial中,计算机程序在屏幕中心呈现一个名词,持续时间为1s。在trail之间,在屏幕中央呈现一个“十”字,以控制被试的注意。120个名词按照伪随机顺序出现,两类刺激交替出现,每完成60个trial后,被试休息3 min。实验完成后,实验者要求被试再次填写状态焦虑量表作为后测水平。
图1 被试声音判断动物和植物分类时的声音波形图Fig.1 Voice waveforms in pronunciations of animal vs plant nouns
实验包括2种反应任务,一种是声音反应任务:实验者要求被试判断在屏幕上呈现的名词的类型(动物或植物),并大声说出该名词的分类(动物或植物)。本研究使用话筒记录被试的声音反应,并从中计算被试的反应时间。为准确记录反应时间,我们编写刺激程序,使其在每一个视觉刺激开始时发出一个短暂的1 000 hz声音(beep)。我们利用Matlab(Mathworks Inc,USA)编写的程序对记录的声音信息首先进行滤波和降噪处理,再计算刺激呈现时(beep出现时)到被试开始发声的时间。一个典型的记录详见图1。第2种是按键反应任务:实验者要求被试判断在屏幕上呈现的名词的类型(动物或植物),并使用游戏控制杆上的两个按键尽快做出反应。整个实验分为2个组块。在一个组块中,出现动物名词按左手键,植物名词按右手键。另外一个组块则相反,出现动物名词按右手键,植物名词按左手键。
1.5 统计学方法
使用Sigmastat3.5软件(SigmaStat Inc.USA)进行统计分析。应用配对样本t检验,首先检验被试在实验前和实验后的SA得分是否有显著变化。在声音反应任务中,分别统计每名被试对判断动植物名词的平均正确率和反应时,然后在组水平(被试间)检验判断动物名词和判断植物名字的正确率和反应时差异(配对样本t检验)。在按键任务中,应用重复测量方差分析(AVOVA)检验刺激分类(动物和植物)与按键手(左手和右手)两个因素对反应时的主效应及交互作用。本研究中计量数据用均数±标准差(±s)表示,以P<0.05为差异有统计学意义。
2 结果
2.1 被试特征
爱丁伯格利手量表的平均分是78,提示所有被试均为右利手。焦虑量表分数的均值为:SA(前测)=32.76,SA(后测)=33.96,TA=36.12。所有的分数均在正常情感状态范围内。状态焦虑分数的2次测量间差异无统计学意义(P=0.430)。
2.2 分类判断的声音反应任务
正确率:与预期一致,本研究实验任务相对简单,动物名词分类的准确率为98% ±1%,植物名词分类的准确率为99% ±1%。配对t检验结果显示,被试对动物和植物的判断正确率差异无统计学意义(P=0.690)。
反应时:在分类判断正确的词语中,被试对动物名词的反应时间为(623±59)ms,对植物名词的反应时间为(661±60)ms。配对t检验结果显示,被试对动物名词的正确判断反应时显著低于对植物名词的正确判断反应时[t(24)=-3.77,P <0.001,图2]。
2.3 分类判断的按键反应任务
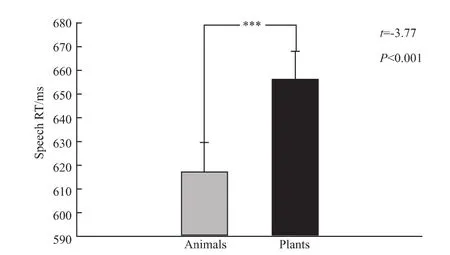
图2 动植物名词声音分类速度比较Fig.2 Categorization speed of animal nouns vs plant nouns with speech response***P <0.001.
与声音反应任务一致,无论使用左手还是右手,被试对动植物名词的判断正确率都接近100%,并且二者差异无统计学意义(左手:P=0.70,右手:P=0.59)。对按键任务反应时的重复测量方差分析(分类×左右手)显示:刺激分类(动物、植物名词)的主效应显著[F1,96)=4.23,P<0.001],左右手的效应不显著[F1,96)=0.68,P>0.05],刺激分类和左右手之间的交互作用显著[F(1,96)=4.42,P<0.001]。事后检验显示,用右手反应,动物名词的反应时(531±13)ms比植物名词的反应时(569±12)ms显著短[t(24)=-3.98,P<0.001]。使用左手反应,动植物名词反应时差异无统计学意义(P=0.31)。左手对动物名词的反应时显著长于右手[t(24)=1.80,P=0.042],左手对植物名词的反应时显著短于右手[t(24)=-1.97,P=0.03](图3)。
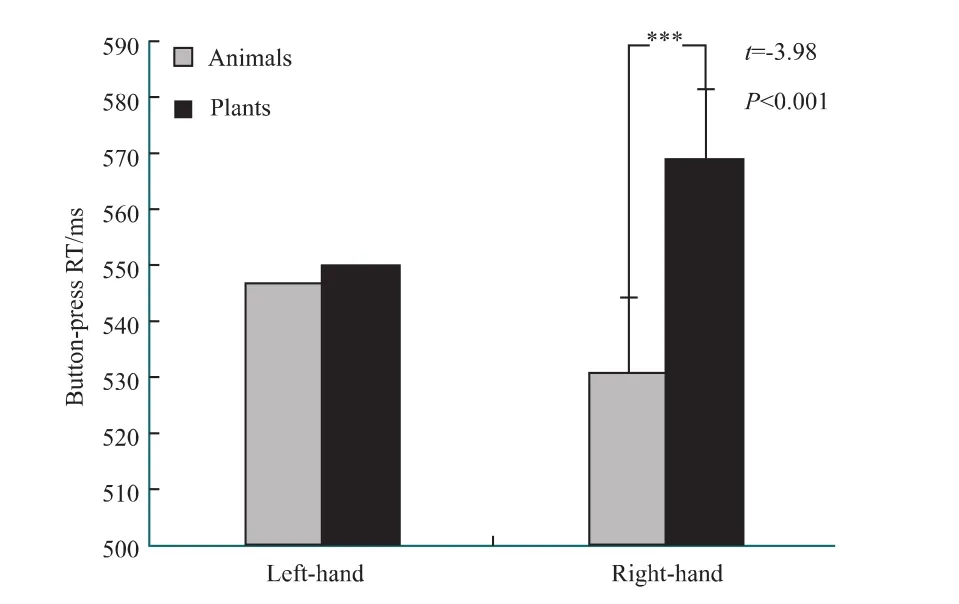
图3 按键反应完成的分类任务中,左右手的比较Fig.3 Comparison of hand effects in categorization task by button-press response***P <0.001.
3 讨论
本研究的结果可总结为:①在正确率接近100%的情况下,被试在声音反应任务中对动物做出判断的反应时显著短于对植物做出判断的反应时;②在按键反应任务中,被试对动植物做出判断的反应时差异受到左/右手的调制。右手按键对动物做出判断的反应时显著低于对植物做出判断的反应时。
在声音反应任务中,被试对动物名词的反应时差异提示人脑对动植物概念的加工机制可能差异有统计学意义。动物名词的加工速度显著快于植物名词,其原因可能源自动植物在人类进化的环境中所起的作用不同:相比于植物,动物对人类生存的威胁更加具有主动性和攻击性,以致人脑需要区别对这两种事物的认知,进而进化为有差别的加工机制。这一原理与其他生物进化出对运动物体的高效加工具有相似性[14-15]。
本研究进一步发现被试在用右手(利手)完成反应的条件下,判断动植物名词的反应时差异有统计学意义,而这一差异在用左手(非利手)完成反应的条件下差异无统计学意义。这一结果说明被试对动植物的反应速度差异受到使用左/右手做出反应的调制,进而提示人脑对动植物概念的加工机制差异可能与人脑的功能偏侧化有关[16-17]。大脑左半球通常是右利手被试的优势脑,也是人类的语言认知加工的优势脑[18-19]。本研究所获结果的一种解释是,对动植物的语言加工差异主要体现了具有语言加工优势的大脑左半球,因而这种差异只有在被试用右手完成反应时才能观测到。此外,本实验发现对动物名词的反应右手快于左手,而对植物名词的反应左手反而快于右手。这也说明大脑功能的偏侧化调制了对动物、植物名词的加工,支持以上的解释。然而脑功能的偏侧化对动植物概念加工过程的调制机制尚待进一步研究。
近期有研究者[20]认识到研究不同分类的认知加工差异对了解人类脑功能特性具有重要意义。他们把动物分为灵长类、禽类、昆虫类,分别研究人脑对它们的视觉加工机制。本研究首次分离动物和植物类别,并提供了人脑对它们具有不同加工机制的初步证据。同时,本研究在按键任务中观测到的偏侧化现象进一步解释了在发声任务中观测到的语言加工优势现象。在发声任务中观测到的结果仅能说明动物名词相对于植物名词的加工优势,但是未能说明这种加工优势的机制。在进一步的按键任务中,这一语言加工优势只体现于利手,说明脑功能的偏侧化与动物名词的加工优势有关。这为今后对这一现象的认知神经科学和心理学研究指明了方向。
本研究全部被试均为女性,因此尚未验证性别对动植物加工机制的影响。此外,儿童的语言加工能力和左右半球的功能分化会随年龄增长而发展,研究这些脑机制的发展和分化对动植物分类加工的影响将使我们更深入了解它们的认知加工机制。因此,未来研究将以不同年龄段的儿童为研究对象,观察他们对动植物概念的加工特点及其动态变化。
[1]Maddox W T,Filoteo J V.Stimulus range and discontinuity effects on information-integration category learning and generalization[J].Atten Percept Psychophys,2011,73(4):1279-1295.
[2]Sartori G,Gnoato F,Mariani I,et al.Semantic relevance,domain specificity and the sensory/functional theory of category-specificity[J].Neuropsychologia,2007,45(5):966-976.
[3]Hemond C C,Kanwisher N G,Op de Beeck H P.A preference for contralateral stimuli in human object-and face-selective cortex[J].PLoS One,2007,2(6):574.
[4]Bai H M,Jiang T,Wang W M,et al.Functional MRI mapping of category-specific sites associated with naming of famous faces,animals and man-made objects[J].Neurosci Bull,2011,27(5):307-318.
[5]Anzellotti S,Mahon B Z,Schwarzbach J,et al.Differential activity for animals and manipulable objects in the anterior temporal lobes[J].J Cogn Neuro Sci,2011,23(8):2059-2067.
[6]Rhodes M,Gelman S A.Five-year-olds'beliefs about the discreteness of category boundaries for animals and artifacts[J].Psychon Bull Rev,2009,16(5):920-924.
[7]Marinovic W,Plooy A M,Arnold D,H.The influence of visual motion on interceptive actions and perception[J].Vision Res,2012,60:73-78.
[8]Hauk O,Pulvermüller F.The lateralization of motor cortex activation to action-words[J].Front Hum Neurosci,2011,5:149.
[9]Bracco P,Anastasi G,Piancino M G,et al.Hemispheric prevalence during chewing in normal right-handed and lefthanded subjects:a functional magnetic resonance imaging preliminary study[J].Cranio,2010,28(2):114-121.
[10]Oldfield R C.The assessment and analysis of handedness:the Edinburgh inventory[J].Neuropsychologia,1971,9(1):97-113.
[11]Spiellberger D C,Gorsuch R L,Lushene R,et al.Manual for the State-Trait Anxiety Inventory(Form Y)[M].PaloAho, California: Consulting Psychologists Press,1983:577.
[12]Zung W W.A rating instrument for anxiety disorders[J].Psychosomatics,1971,12(6):371-379.
[13]北京语言学院语言教学研究所.现代汉语频率词典[M].北京:北京语言学院出版社,1985.
[14]Kanakogi Y,Itakura S.Developmental correspondence between action prediction and motor ability in early infancy[J].Nat Commun,2011,2:341.
[15]Machado S,Cunha M,Portella C E,et al.The role played by the parietooccipital cortex in the process of sensory-motor integration:an electroencephalographic study[J].Rev Neurol,2008,47(3):146-149.
[16]汤艳清,欧凤荣,吴枫,等.中美两国6种基本面部表情识别的跨文化研究[J].中国医科大学学报,2011,40(5):422-424,427.
[17]李华,赵文新,任苏虹,等.脑小血管病与轻度认知功能障碍的关系[J]. 中国脑血管病杂志,2009,6(2):70-73.
[18]Nagae L M,Zarnow D M,Blaskey L,et al.Elevated mean diffusivity in the left hemisphere superior longitudinal fasciculus in autism spectrum disorders increases with more profound language impairment[J].AJNR Am J Neuroradiol,2012,Apr 5.[Epub ahead of print]
[19]Jones G V,Martin M.Language dominance,handedness and sex:recessive X-linkage theory and test[J].Cortex,2010,46(6):781-786.
[20]Connolly A C,Guntupalli J S,Gors J,et al.The representation of biological classes in the human brain[J].J Neurosci,2012,22,32(8):2608-2618.
