崇明东滩斑背大尾莺繁殖生态分析1)
2012-08-29丛日杰胡春芳东北林业大学哈尔滨150040汤臣栋庚志忠冯雪松崇明东滩鸟类国家级自然保护区张玉铭东北林业大学
丛日杰 李 枫 胡春芳(东北林业大学,哈尔滨,150040)汤臣栋 庚志忠 冯雪松(崇明东滩鸟类国家级自然保护区)张玉铭(东北林业大学)
斑背大尾莺(Locustella pryeri)隶属于雀形目(Passeriformes)莺科(Sylviidae)蝗莺属(Locustella)[1]。已有研究表明[2-7],该鸟在国内主要有4个繁殖地,分别是黑龙江扎龙、辽宁双台河口、江西南矶山及上海崇明东滩。在崇明东滩地区斑背大尾莺为四季常见鸟类,主要在芦苇和互花米草群落中栖息营巢。斑背大尾莺有两个亚种,分别繁殖于日本(指名亚种)和中国东北[8](汉口亚种),在我国主要为汉口亚种。
目前关于斑背大尾莺的研究主要集中在种群分布[6-7,9-12]、鸣唱[2]、遗传结构[13]、繁殖生态[5]等方面。笔者于2010年、2011年4—6月在上海崇明东滩鸟类国家级自然保护区对其繁殖生态习性进行了研究,以了解斑背大尾莺不同地理种群间繁殖生态是否会出现分化。
1 研究地概况
崇明东滩位于崇明岛的东端,地理位置在N31°25'~31°38',E121°50'~122°05'之间,是由长江携带的泥沙沉积而形成的滩涂,为长江口规模最大、发育最完善的河口型潮汐滩涂湿地[14]。受北亚热带海洋气候影响,气候温和,四季分明。年均气温15.3℃,年均降水量为1 022 mm,相对湿度为82%[15]。东滩湿地由于受到潮汐、淹水以及盐度胁迫等的影响,生长的植物种类很少,植被类型单一,主要由海三棱藨草(Scirpus mariqueter)、芦苇(phagmites cemminis)和藨草(Scripus triqueter)这3大本地滩涂群落类型与互花米草(Spartina alterniflora)群落组成。其中互花米草为外来入侵种,通过人为引入和自然扩散在崇明东滩的滩涂湿地迅速扩展,排挤当地土著种芦苇(phagmites cemminis)和海三棱蔗草(Scirpus mariqueter),改变了植物群落结构,导致鸟类多样性下降[16]。
2 研究方法
分别于2010年和2011年的4月至6月间,借助10倍望远镜在崇明东滩对斑背大尾莺繁殖情况进行了监测,观察该鸟的占区、求偶、筑巢、产卵、育雏等与繁殖相关的行为特征,根据其行为特征寻找鸟巢;并随机选择巢进行卵大小(长径、短径)、卵质量及巢参数的测量和统计。同时采用温湿计对亲鸟离巢后的孵化温度进行全日制测量。育雏期每日同一时间对雏鸟的生长发育情况进行记录,主要参数包括:体质量、体长、翅长、跗跖长和尾长等。
产卵期以各巢产首枚卵的日期作为指标,一般根据卵孵化的日期倒推而得或根据卵失重的数值进行估算[17]。雏鸟体质量以及部分外部器官的生长用Logistic曲线拟合,渐进线由三点法求出,并求出与拟合程度相关的系数R2[5]。使用Krsukal-Wallis检验对崇明东滩、双台河口、扎龙3地巢参数、卵参数的差异进行比较,所有统计分析均在SPSS18.0上完成,文中数据均以Mean±SD的形式表示。
3 结果与分析
3.1 筑巢与巢
斑背大尾莺3月末占区,4月中旬开始筑巢。据对19个巢的观察,筑巢时间为6 d左右。筑巢主要由雌鸟完成,雄鸟不参与筑巢。
斑背大尾莺巢十分隐蔽,筑在枯互花米草的底部,巢距地面有一定的距离;巢材为枯互花米草和芦苇叶,内垫物常为细软的苔草或成鸟羽毛等。新筑的巢分为椭圆状有盖和浅杯状无盖两种巢型。巢口直上直下或略微倾斜,多被倒伏的互花米草覆盖。随着亲鸟进出巢频次的增加,有盖巢的巢口逐渐扩大,到繁殖期结束时,巢型区分不明显,两种巢型均接近碗状。19个巢的参数为:巢的内径平均值(64.95±10.16)mm,外径(100.63±10.64)mm,巢高(98.32±15.84)mm,巢深(68.42±9.03)mm,巢距地面距离(182.21±99.9)mm。
3.2 产卵
斑背大尾莺每年产卵高峰期是4月下旬和5月上旬,产卵时间持续到5月末。产卵期3~6 d,日产卵1枚。鲜卵为透明浅粉色,孵化后期呈乳白色。鲜卵质量(1.43±0.15)g(n=20枚),长径(17.85±1.24)mm(n=20枚),短径(12.43±1.47)mm(n=20枚),窝卵数(4.59±1.2)枚(3~6枚,n=17巢)。
对2010—2011年该鸟窝卵数频次进行统计(图1,n=17巢),结果表明:窝卵数为5枚的巢最多,占巢总数的35.29%,所以5枚为该鸟常见窝卵数。在产卵后期,人为将卵移走1~2枚,巢内剩余卵数仍保持不变,说明该鸟不具补卵行为。

图1 窝卵数频次
3.3 孵化
产完卵后由雌鸟孵化,孵化期10~12 d(n=4)。孵化期巢内每日平均温度:(28.35±0.28)℃(28.36~29.39℃,n=11 d)(如表1),温度波动幅度在1℃左右。5:00—18:00巢内温度随时间变化如表2所示。1 d中巢内孵化温度的范围:23.85~32.95℃,且最小值和最大值分别出现在5:00和13:00—14:00时间段,因此推测孵化温度受外界气温影响较大。孵化期,笔者对孵化期卵的失重情况(表3)进行了统计,通过失重率公式[18],即失重率=(鲜卵质量-出雏前卵质量)/鲜卵质量×100%,得出其卵的平均失重率为21.98%(n=10枚)。

表1 斑背大尾莺巢内孵化温度的日变化
3.4 育雏及雏鸟的生长发育
雌雄共同育雏,育雏期14 d左右,包括巢内育雏和巢外育雏两阶段。巢内育雏9~10 d(平均值=(9.18±0.41)d,n=11巢),该阶段雏鸟生长发育较快,完全依靠亲鸟喂食。巢外育雏4~5 d(平均值=(4.75±0.45)d,n=11巢),雏鸟虽已离巢但其羽毛尚未丰满,飞翔及捕食能力较弱,仍需亲鸟喂食。
雏鸟外部器官生长方程如表4所示。体质量、翅长、跗跖生长遵循logistic方程规律。其中:体质量、附跖在第4~5天生长最快;而翅长的拐点较小,说明其生长发育较迅速。体长、嘴长均遵循线性生长规律。尾羽于第5天长出,呈指数形式增长,其生长公式:y=e0.42t-1.41,R2=0.998。
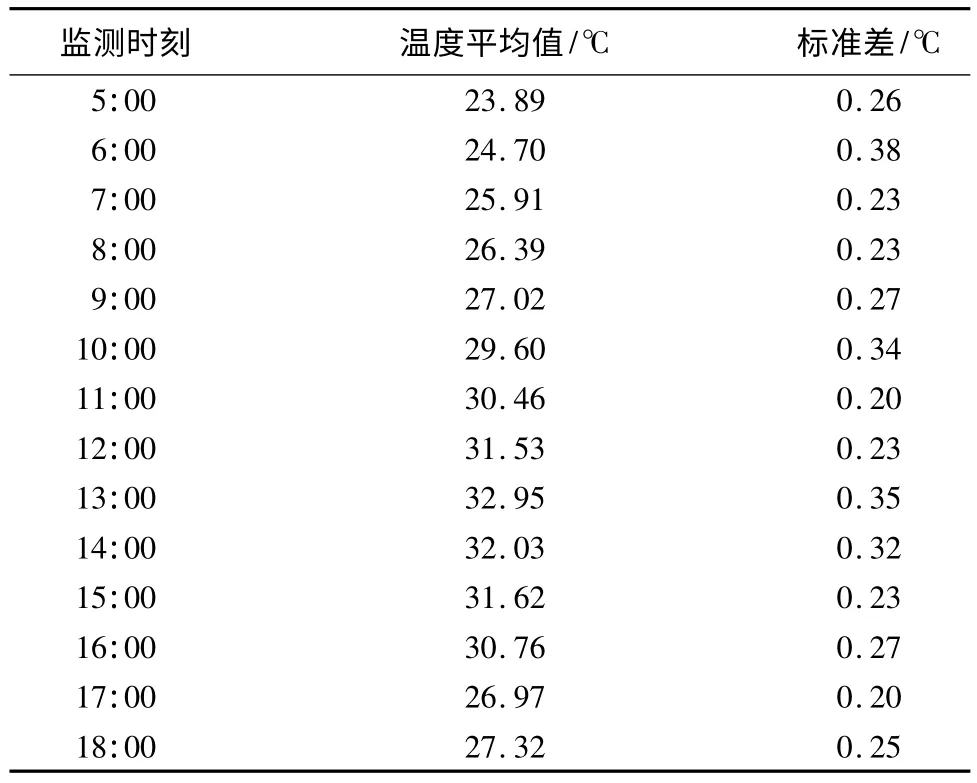
表2 斑背大尾莺巢内孵化温度的变化

表3 斑背大尾莺孵化期卵质量损失情况

表4 雏鸟生长方程
3.5 繁殖成效
2010—2011年斑背大尾莺的繁殖结果如表5所示。2011年的繁殖情况与2010年相比,其孵化成功率、卵成功率均有所上升。
3.6 繁殖失败原因
两年中有10巢繁殖失败,其中5个巢在孵化期被天敌捕食;3个巢在育雏期被天敌捕食;2个巢由于人为干扰而弃巢。78枚卵中有36枚孵化失败,其中22枚卵在孵化期被捕食,9枚卵死胎,5枚卵未受精。因此崇明东滩地区斑背大尾莺巢捕食、干扰、卵未受精和死胎是导致繁殖失败的主要因素。

表5 繁殖结果
3.7 繁殖参数比较
对扎龙、双台河口、崇明三地斑背大尾莺的繁殖参数进行独立样本Krsukal-Wallis检验,3地的卵参数差异显著(卵长径差异不显著),如表6所示。斑背大尾莺在崇明东滩地区的巢型和巢参数与其它两地有较大区别,在崇明东滩巢型主要以浅杯状无盖巢为主,而其他两地则主要以椭圆状有盖巢为主。此外巢高、巢深、巢外径、巢内径等参数与其它两地差异显著,如表6所示,这种不同地区巢参数差异较大的现象与乌鸫类似[20]。

表6 3地斑背大尾莺繁殖参数Krsukal-Wallis检验
4 结论与讨论
窝卵数代表了子代质与量之间的权衡[21],卵的大小不仅与食物可利用性有关[22],还可受温度[23]、海拔[24-25]等多种因素的影响,因此3个繁殖地卵参数差异是环境因素、遗传因素和繁殖策略3者综合作用的结果。斑背大尾莺在不同地区可利用的巢材各异,其巢参数差异也较大。因而,可利用巢材的不同也可能是斑背大尾莺在不同地区巢参数差异的原因。但是这种可能性以及是否存在其它原因还有待以后的工作进一步核实。
巢址周围地域的植被结构特征影响着巢卵被捕食率[26]。崇明东滩地区巢型与其它地区差异较大,主要因为扎龙、双台河口、崇明3地巢址选择机制的不同。对于崇明东滩地区出现的两种巢型,笔者推测是该鸟在崇明东滩地区生存时间较短,营巢策略进化不稳定的表现。此外,Slagsvold[27]认为,巢尺度的减小是由窝卵数的减小所导致。崇明东滩巢尺度及窝卵数与扎龙相比明显减少,支持上述研究结果。
崇明东滩繁殖种群与其他两个种群相比在繁殖生态方面有一定的分化,导致这种分化是由于3者的繁殖策略差异还是种内分化所致,仍有待在生活史方面及分子水平上进行深入研究。
[1]郑作新.中国鸟类区系纲要[M].北京:科学出版社,1987:779-780.
[2]曲文慧,李枫,沙剑斌,等.斑背大尾莺繁殖期鸣声行为分析[J].动物学研究,2011,32(2):141-149.
[3]何芬奇,林剑声,黄小江,等.斑背大尾莺鄱阳湖繁殖亚群初报[J].动物学杂志,2008,43(2):70-72.
[4]郑光美.中国鸟类分类与分布名录[M].北京:科学出版社,2005:294-317.
[5]王强,李枫.斑背大尾莺sinensis亚种的繁殖生物学[J].动物学报,2006,52(6):1162-1168.
[6]鲁长虎,李枫.黑龙江省鸟类新纪录:斑背大尾莺[J].四川动物,1997,16(3):104.
[7]干晓静,章克家,唐仕敏,等.上海地区鸟类新记录3种:史氏蝗莺、斑背大尾莺、钝翅苇莺[J].复旦学报:自然科学版,2006,45(3):417-420.
[8]约翰·马敬能,卡伦·菲利普斯,何芬奇.中国鸟类野外手册[M].卢和芬,译.长沙:湖南教育出版社,2000:361-362.
[9]郑作新.中国鸟类种和亚种分类名录大全[M].北京:科学出版社,2000:107-117.
[10]高明,殷秀琴.斑背大尾莺的白色型[J].动物学杂志,2007,42(6):20-21.
[11]郑光美.中国鸟类分类与分布名录[M].北京:科学出版社,2005:294-317.
[12]何芬奇,林剑声,黄小江,等.斑背大尾莺鄱阳湖繁殖亚群初报[J].动物学杂志,2008,43(2):70-72.
[13]Zhang Weiwei,Zhang Yuming,Wang Qiang,et al.Genetic diversity and population structure of the Marsh Grassbird(Locustella pryeri sinensis)in China[J].Journal of Forestry Research,2010,21(3):361-366.
[14]徐宏发,赵云龙.上海市崇明东滩鸟类自然保护区科学考察集[M].北京:中国林业出版社,2005.
[15]黄华梅,张利权,袁琳.崇明东滩自然保护区盐沼植被的时空动态[J].生态学报,2005,27(10):4166-4172.
[16]Chen Zhongyi,Li Bo,Zhong Yang,et al.Local competitive effects of introduced Spartina alterniflora on Scirpus mariqueter at Dongtan of Chongming Island,the Yangtze River estuary and their potential ecological consequences[J].Hydrobiologia,2004,528(1/3):99-106.
[17]张正旺,郑光美,杨向明,等.褐马鸡窝卵数及其变异[J].动物学杂志,1997,43(增刊1):83-86.
[18]孙森,刘志平,叶春艳.野禽饲养学[M].哈尔滨:东北林业大学出版社,1995:73.
[19]Bond R R.Ecology distribution of breeding birds in the upland forests of southern Wisconsin[J].Ecol.Monogr,1957,27(4):351-384.
[20]黄族豪,程亚林,梅文枫,等.江西吉安乌鸫繁殖生态研究[J].四川动物,2011,30(3):439-441.
[21]赵亮,张晓爱,李明才.窝雏数处理对两种雀形目幼鸟生长的影响[J].动物学杂志,2002,37(3):6-8.
[22]Christians J K.Avian egg size:variation within species and inflexibility within individuals[J].Biological Reviews,2002,77(1):1-26.
[23]Nager R G,Van Noordwijk A J.Energetic limitation in the egglaying period of great tits[J].Biological Sciences,1992,294:259-263.
[24]Milinkovich D J.The sources of variation in the reproductive characters of House Wrens(troglodytes aedon)breeding at two elevations in Colorado[D].Tallahassee,FL:Florida state university,1993.
[25]Johnson L S,Ostlind E,Brubaker J L,et al.Changes in egg size and clutch with elevation in a Wyoming population of Mountain Bluebirds[J].The Condor,2006,108(3):591-600.
[26]Martin TE,RoPer JJ.Nest predation and nest-site selection of a western population of the Hermit Thrush[J].The Condor,1998,90(1):51-57.
[27]Slagsvold T.On the evolution of clutch size and nest size in passerine birds[J].Oecologia,1989,79(3):300-305.
