德国镜鲤和不同倍性泥鳅银染核型的比较研究
2012-08-08李雅娟曹顶臣孙效文
李雅娟,高 敏,钱 聪,曹顶臣,孙效文
(1.大连海洋大学农业部北方海水增养殖重点实验室,辽宁 大连 116023;2.中国水产科学研究院黑龙江水产研究所,哈尔滨 150070)
德国镜鲤(Cyprinus carpioL.mirror)具有生长速度快、含肉率高、肉质好、易捕捞等优点,它与黑龙江野鲤、红鲤等杂交,其杂种后代都表现出明显的杂种优势[1]。泥鳅(Misgurnus anguillicaudatus)是我国常见的一种小型经济鱼类。泥鳅肉质鲜美,营养丰富,富含蛋白质及多种维生素,并具有药用价值[2]。泥鳅存在多倍体现象,我国除了二倍体泥鳅外还存在天然三倍体、四倍体泥鳅[3-7]。最近Li等研究表明,我国自然四倍体泥鳅是含有四套染色体组的遗传四倍体,是多倍体育种的良好材料[8-9]。核仁组织区(Nucleolus organizer region,NOR)是真核细胞染色体上18s+28s r RNA基因所在的部位,是生产rRNA的场所。用硝酸银染色法可显示有活性的Ag-NOR。NOR数目被认为是一个稳定的细胞遗传学指标。用携带有Ag-NORs的染色体作为鱼类核型进化的指标,对于探讨鱼类系统发生具有重要意义。这种方法已在哺乳类和两栖类等脊椎动物获得广泛应用。自Klingerman等最早对鱼类NOR进行分析以来,这方面的研究逐渐增多[10]。据不完全统计,迄今已对200余种鱼的染色体NOR进行了分析。国内对于鱼类银染带的研究起步比较晚,中国具有世界上最为丰富的鲤科鱼类[11],关于鲤科鱼染色体核仁组织区(NOR)研究也曾有报道。桂建芳等对四种鲤科鱼类的银染核型进行了研究[12]。王蕊芳等对鲤鱼的一些品种的染色体银染带及不同地理区域鲫鱼染色体银染核仁组织者的进行了比较研究[14]。任修海等对36种鲤科鱼染色体核仁组织区(NOR)研究指出,中国鲤科鱼类NOR的基本特征是,2对NOR存在于亚中部着丝粒染色体上,属于较特化的类型[15]。杨太有等对黄河鲤的染色体核型及银染核型进行了研究[16]。鲤科鱼类的倍性问题一直是科学家的研究热点,据报道德国镜鲤染色体数目为100[17],它是具有4套染色体组的遗传的四倍体(4n=100),还是已经二倍化的进化的四倍体(2n=100),目前尚未清楚。因此,本文通过对德国镜鲤与二倍体泥鳅、四倍体泥鳅的银染核型进行比较分析,旨在为德国镜鲤和泥鳅的起源提供细胞遗传学证据,并为其遗传、变异、分类、系统演化以及杂交育种提供参考。
1 材料与方法
1.1 实验用鱼的采集
德国镜鲤采自黑龙江省呼兰永兴鱼场(雌鱼22尾,雄鱼10尾),暂养于实验室水族箱,暂养温度为(22±2)℃。泥鳅采自长江水系湖北省赤壁市,运回实验室暂养,暂养温度(23±1)℃。
1.2 染色体标本的制备
每尾鱼腹腔注射PHA溶液,剂量为6 μg·g-1(鱼体重),18~20 h后进行第二次注射PHA(剂量同第一次),作用4~6 h后,活体腹腔注射0.1%秋水仙素,剂量为6 μg·g-1(鱼体重),作用2~3 h。剪尾鳍放血5~10 min。分别取鳃、肾组织剪成小块,置于0.8%的柠檬酸钠溶液中,室温低渗40~45 min。再将鳃、肾组织块移入新配制预冷的卡诺固定液(甲醇∶冰醋酸=3∶1)中,每隔15 min更换一次固定液,重复3次。加入新的固定液置-20℃冰箱中过夜保存,次日取部分组织块用50%冰醋酸解离,剪成细胞悬液,加入新配置卡诺固定液,过滤。采用冷滴片法制备染色体标本。
1.3 银染(Ag-NOR)
采用Howell和Black[18]的快速银染法。50%硝酸银与2%明胶(内含1%甲酸)以2∶1混匀后滴在染色体标本上,加盖玻片,70℃处理2 min。当整张玻片呈棕黄色时取出,流水冲去盖玻片,干燥后镜检。
1.4 核型分析方法
分别观察50个分裂相和100个细胞核,以确定Ag-NORs的数目。选取3~5个清晰、分散较好的分裂相进行显微拍照,并分别进行染色体计数。核型分析参照文献[19]的方法进行,以确定Ag-NORs的位置。
2 结果与分析
2.1 Ag-NORs的数目
Ag-NORs的数目在中期分裂相及间期核中均表现出不同的多态性。德国镜鲤中期分裂相及间期核中呈现出Ag-NORs的数目为1~2个,含有2个Ag-NORs的频率最高,分别为80%、56%(见表1,图1-A)。二倍体泥鳅中期分裂相及间期核中呈现出Ag-NORs的数目为1~2个,含有2个Ag-NORs的频率最高,分别为72%、60%(见表1,图1-B)。四倍体泥鳅中期分裂相及间期核中呈现出Ag-NORs的数目为1~4个,中期分裂相中含有3个Ag-NORs的频率最高,为36%(图1-C),间期核中含有2个Ag-NORs的频率最高,为40%(图1-C,图1彩版见封三)。
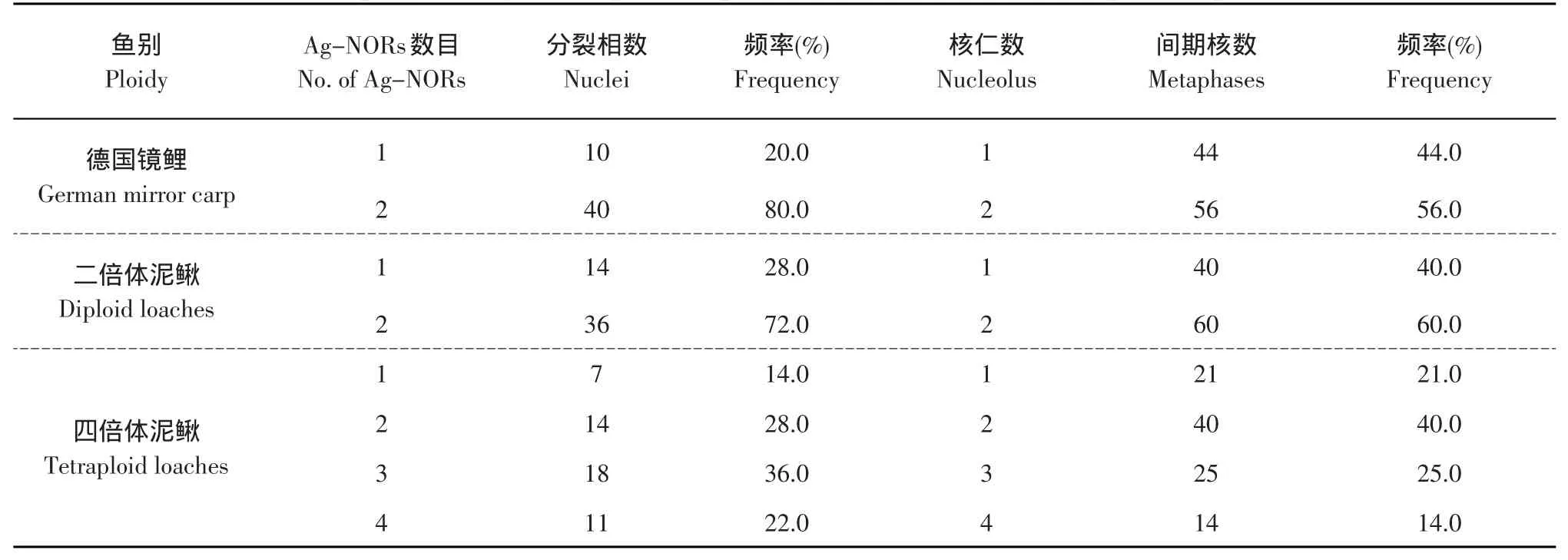
表1 中期分裂相Ag-NORs频率及间期核中核仁的频率Table 1 Frequency of Ag-NORs per metaphase and number of nucleolus per interphase nucleu
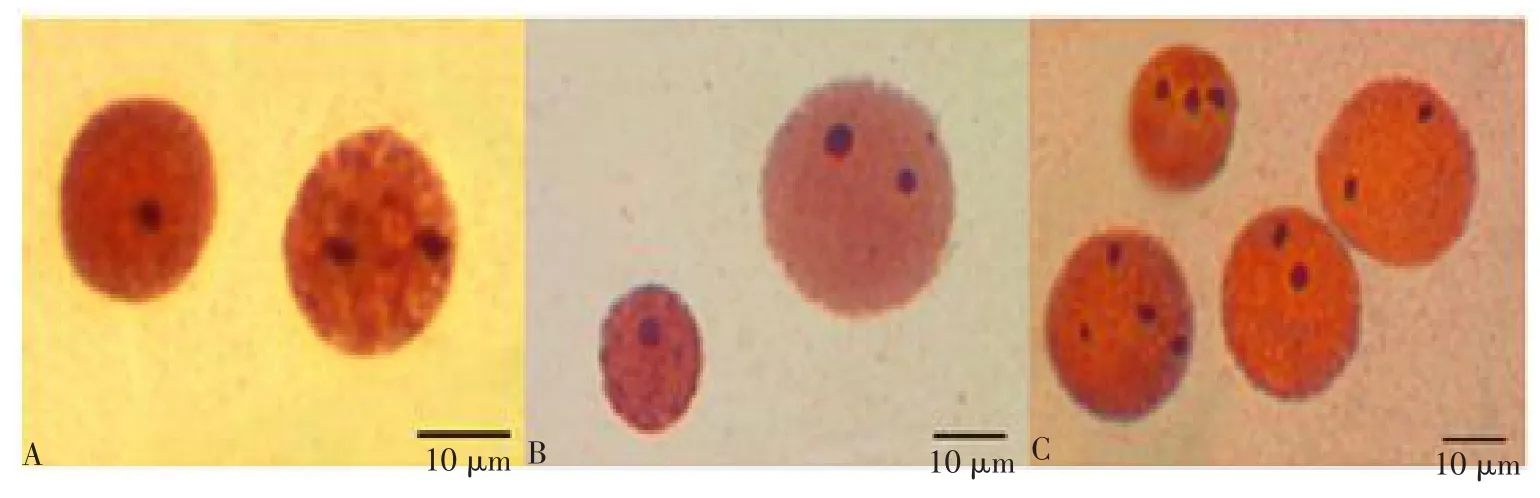
图1 德国镜鲤和不同倍性泥鳅间期核硝酸银染色Fig.1 Ag-NORs in interphase nuclei from German mirror carp and different ploidy loaches
2.2 Ag-NORs的核型
为了确定携带有NOR的染色体在核型中的位置和NORs在染色体上的位置,对德国镜鲤和不同倍性泥鳅进行银染核型分析。
2.2.1 德国镜鲤
2n=100,核型组成为30 m+26 sm+30 st+14 t,NF=156。NOR的染色体在核型中位置和NORs在染色体上的位置是:sm组第8号染色体的短臂末端(见图2-A,D,图2彩版见封三)。
2.2.2 二倍体泥鳅
2n=50,核型组成为10 m+4 sm+36 st,NF=64。NOR的染色体在核型中位置和NORs在染色体上的位置是:m组第1号染色体的短臂末端(见图2-B,E)。
2.2.3 四倍体泥鳅
4n=100,核型组成为20 m+8 sm+72 st,NF=128。NOR的染色体在核型中位置和NORs在染色体上的位置是:m组第1号染色体的短臂末端(见图2-C,F)。

图2 德国镜鲤和不同倍性泥鳅银染中期分裂相及核型Fig.2 Ag-NORs and Karyotypes in metaphase spreads from German mirror carp and different ploidy loaches
3 讨论与结论
3.1 德国镜鲤与鲤科鱼类银染核型的比较
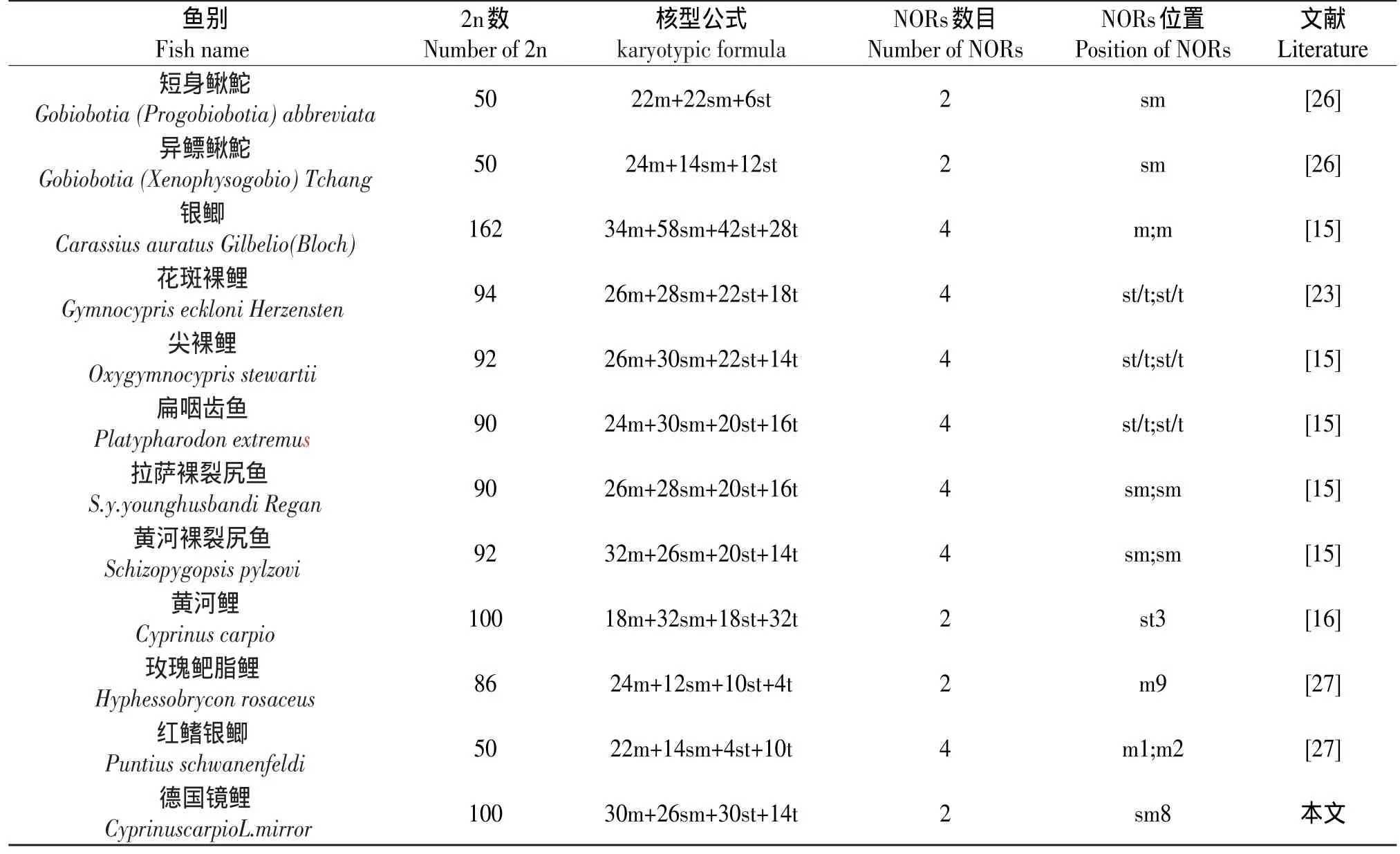
据不完全统计,迄今国内已对50余种鲤科鱼的Ag-NORs进行分析(见表2),从表2中可以看出,具有4个Ag-NORs的种类占59%;其次是具有2个Ag-NORs占25%;具有6个Ag-NORs占10%;具有8个Ag-NORs占6%。Ag-NORs染色体的类型是:sm占51%;st占25%;t占13%;m占11%。
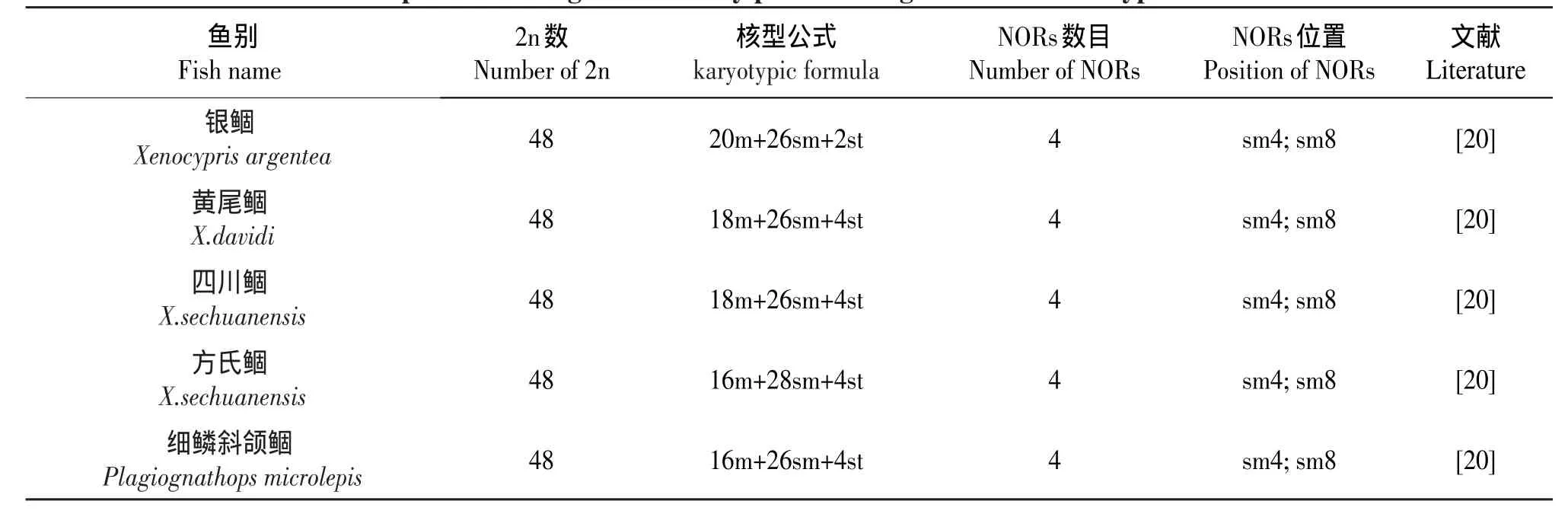
表2 目前国内已发表的鲤科鱼Ag-NORs资料比较Table 2 The comparison among the currently published Ag-NORs data of Cyprinid fishes in China
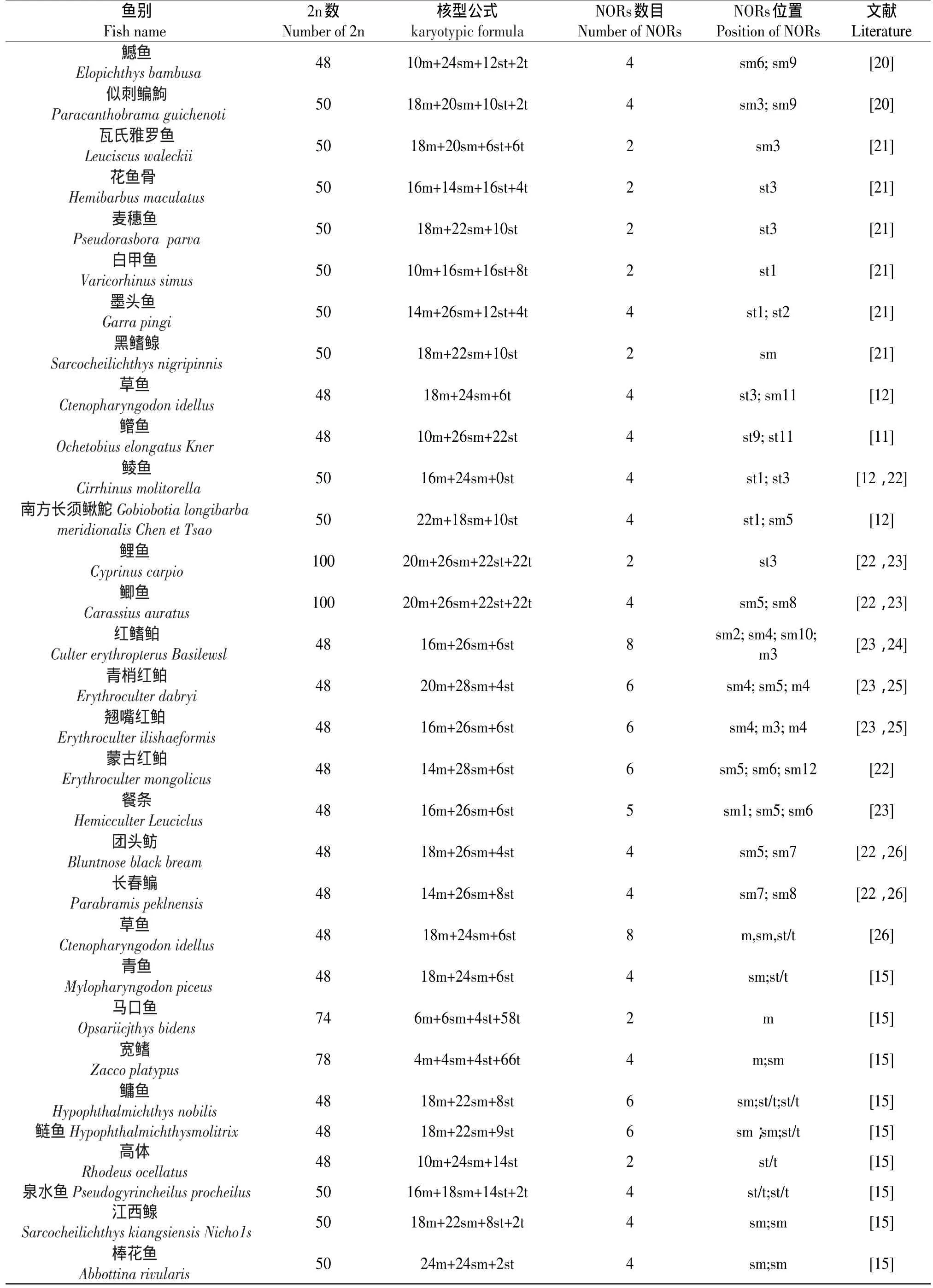
续 表
续 表
从表2所列举的银染资料可以看出,多数中国鲤科鱼类的Ag-NORs数为4,位于两对亚中部着丝点染色体的短臂末端,说明中国鲤科鱼类具有较特殊的银染核型。Amwemiya和Gold[29]等通过对50余种北美鲤科鱼类研究发现,绝大多数都具有一对携带NORs的染色体,并指出,具有一对NORs染色体的类型为鲤科鱼类的原始类型。Takai和Ojima根据对日本鲤科鱼的分析也认为,具有一对较小的NOR是鱼类中较基本和原始的性状[29]。Glod等根据“普遍性原则”推测亚端部着丝点染色体短臂上的Ag-NORs为鱼类中最原始类型[30]。根据以上分析,可以认为,中国鲤科鱼类的原始NOR特征也应是一对NOR、位于端部着丝粒(st或t)染色体短臂末端。本研究发现,德国镜鲤具有2个Ag-NORs,并位于sm染色体上,说明是原始类型。
3.2 德国镜鲤染色体多倍化
鲤科鱼类基本的染色体数是50或48,这已被大多数学者所公认。Arai[31]在总结分析了欧亚大陆已作过核型研究的141种鲤科鱼类的基础上,认为2n=50是鲤科鱼类最原始的核型,并由2n=50朝几个方向演化,即:1.通过非整倍性增加或着丝粒断裂,由2n=50→52;2.通过着丝粒断裂,由2n=50或48→78;3.通过多倍化增加,由2n=50→100或98;4.通过罗伯逊易位,由2n=50→48,2n=48→44;5.通过缺失而减少,由2n=48→46。在北美鲤科鱼类中90%以上的物种染色体数目2n=50。据周暾[32]统计和咎瑞光[33]报道,在我国已做过染色体研究的117种鲤科鱼类染色体中,2n=50的有49种,占42%,所以众多学者公认50或48条染色体是鲤科鱼类基本的二倍体条数。这些数据说明2n=50的核型是鲤科鱼类较原始的核型[23]。近年来,对鲤科鱼类染色体研究比较多,从已有的报道中发现,染色体数2n=100广泛存在于鲤亚科的鲤属和鲫属。本研究德国镜鲤的染色体数目为100,是50的2倍,这与已作核型分析的中国鲤鱼相同。经银染分析,最高银染点数代表细胞染色体的倍性,四倍体泥鳅具有4个NORs,德国镜鲤和中国鲤鱼均具有2个NORs,分别分布在sm染色体上,与大多数的中国鲤科鱼类NOR表型一致[15]。四倍体泥鳅无论中期分裂相还是间期核最高银染数为4个。因此,认为四倍体泥鳅进行加倍后未发生二倍化或在二倍过程中,而德国镜鲤与中国鲤鱼一样是多倍化后经历了二倍化过程演变而来,是二倍化的四倍体。本研究结果可为德国镜鲤和泥鳅的亲缘关系及起源提供细胞遗传学证据。
[1] 尹洪滨,刘明华,沈俊,孙中武.德国镜鲤的核型研究[J].生物技术,1995,5(3):16-18.
[2] 刘孝华.泥鳅的生物学特性及养殖技术[J].湖北农业科学.2008,47(1):93-95.
[3] 李渝成,李康.马口鱼和泥鳅的核型研究兼论鱼类染色体数目多态与分类的关系[J].武汉大学学报,1987,1:107-112.
[4] 李雅娟,印傑,王嘉博,等.中国29の地点ドジョウにおける倍数体の分布に関する研究[J].日本水産学会誌,2008,74(2):177-182.
[5] 李康,李渝成,周暾.两种泥鳅染色体组型的比较研究[J].动物学研究,1983,4:75-81.
[6] 印杰,赵振山,陈小奇,等.二倍体和四倍体泥鳅染色体组型比较[J].水生生物学报,2005,4(29):469-472.
[7] 李雅娟,田萍萍,李莹,等.中国洪湖不同倍性泥鳅的染色体组型及形态特征比较分析[J].大连水产学院学报,2009,24(3):236-240.
[8] Li Y J,Tian Y,Zhang M Z,et al.Chromosome banding and FISH with an rDNA probe in the diploid and tetraploid LoachMisgurnus anguillicaudatus[J].IchthyologicalResearch,2010,57(4):358-366.
[9] Li Y J,Yu Z,Zhang M Z,et al.The origin of natural tetraploid loach Misgurnus anguillicaudatus(Teleostei:Cobitidae)inferred from meiotic chromosome configurations[J].Genetica,2011(139):805-811.
[10] Klingerman A D,Bloom S E.Distribution of F-bodies,heterochromatin and nucleolar organizers in genome of central mudminnow[J].Umbra limi Cytogenet Cell Gent,1977,18:182-196.
[11] 伍献文编.中国鲤科鱼类志(下卷)[M].北京:科学出版社,1977:229-570.
[12] 桂建芳,周暾.四种鲤科鱼类和一种鮠科鱼类的银染核型研究[J].武汉大学学报,1986(1):106-112.
[13] 王蕊芳,施立明,贺维顺.几种鲤鱼染色体核仁组织者的银染观察[J].动物学研究,1985,6(4):391-398.
[14] 王蕊芳,施立明,贺维顺.不同地理区域鲫鱼染色体银染核仁组织者的比较研究[J].动物学研究,1988,9(2):165-169.
[15] 任修海,崔建勋,余其兴.中国鲤科鱼类染色体核仁组织区研究[J].武汉大学学报:自然科学版,1996,42(4):475-480.
[16] 杨太有,李仲辉,张西瑞,等.黄河鲤鱼核型和银染核型的研究[J].淡水渔业,1996,26(4):6-8.
[17] 张克俭,何玉明,张景龙,等.德国镜鲤染色体组型的研究[J].上海水产大学学报,1999,54(1):72-75.
[18] Howell W,Black M.Controlled silver-staining of nucleolus organizer regions with a protective colloidal developer:A 1-step method[J].Exppeneertia,1980,36:1041-1015.
[19] Levan A,Fredga K,Sandberg A A.Nomenclature for centromeric position on chromosomes[J].Hereditas,1964,52:201-220.
[20] 周密,康杨,李渝成,等.鲤科七种鱼的银染核型研究[J].动物学研究,1988,9(3):225-229.
[21] 任修海,崔建勋,余其兴.六种鲤科鱼类核仁组织者区的研究[J].遗传,1993,15(4):11-13.
[22] 常重杰,余其兴.鲤和鲫银染核型的比较研究[J].武汉大学学报:自然科学版,1993(3):110-114.
[23] 任修海,余其兴,崔建勋,等.鱼类染色体的荧光显带研究[J].遗传学报,1993,20:116-121.
[24] 常重杰,余其兴.七种鲌亚科鱼Ag-NORs的比较研究[J].遗传,1997,19(4):22-25.
[25] Ren Xiuhai,Yu Xianjue.Characterization of nucleolar organizer regions of twelve species of Chinese cyprinid fishes[J].Caryologia,1993,46:201-207.
[26] Ren Xiuhai,Cui Jianxun,Yu Qixing.The heterogeneities of heterochromatins and NORs in the genomes of chinese cyprinid fishes[J].Cytologia,1991,56:673-677.
[27] 陈友铃,郑翠芳,吴文珊,等.玫瑰脂鲤和红鳍银鲫的核型及银染和C-带研究[J].福建师范大学学报:自然科学版,2007,23(4):96-100.
[28] Amemiya C T,Gold J R.Cytogenetic studies in North American minnows(Cyprinidae),Chromosomal NOR phenotypes of 12 species,with comments on cytosystematic relationships among 50 species[J].Hereditas,1990,112:231-247.
[29] Takai A,Ojima Y.Chromosomal distribution of nucleolus organizer in Japanese cyprinid fish[J].Cytobios,1992,71:7-17.
[30] Gold J R.Silver-staining and heteromorphism of chromosomal nucleolus organizer regions in North American cyprinid fishes[J].Copeia,1984(1):133-139.
[31] Arai R.A chromosome study on two cyprinid fishes,A crossochellusis bistus andPteudorabora pumilswith notes on Eurasian cyprinidae and their karyotypes[J].Bull Sci Mus Tokyo Ser,1982,8(3):131-152.
[32] 周墩.鱼类染色体研究[J].动物学研究,1884,5(增刊):38-52.
[33] 咎瑞光,宋峥,刘万国,等.七种鲃亚科鱼类的染色体组型研究[J].兼论鱼类多倍体的判定问题动物学研究,1984,5(增刊):82-90.
