有氧运动对肥胖大鼠脂肪组织perilipin A基因表达的影响
2012-08-06封飞虎杨月琴
傅 姗,封飞虎,杨月琴
(1.温州医学院 体育科学学院,浙江 温州325035;2.武汉体育学院,湖北 武汉430079)
肥胖是由于长期的脂肪组织中三酰甘油的储存与动员之间的不平衡所[1]。虽然脂肪水解的基本规律并未被洞悉,但多个动物实验均已证明,脂肪细胞中的特殊蛋白perilipin在脂解过程中的起了重要作用。Perilipin是一种紧密结合在成熟脂肪细胞脂滴表面的蛋白,中文译名为:脂滴包被蛋白、围脂滴蛋白、脂被蛋白或周脂素。大鼠的perilipin基因生成多个mRNA片段,编码生成至少四种亚型。尽管B亚型也在脂肪细胞中表达,但A亚型是含量最大的。C亚型及D亚型只在甾体细胞中表达[2]。近年来关于脂滴包被蛋白的结构与功能,脂滴包被蛋白的表达与其调控脂解机制及其影响因素,以及脂滴包被蛋白与肥胖以及相关疾病关系的研究成为热点。许多运动医学研究者也开始关注脂滴包被蛋白的研究,但国内外文献相对较少。本研究拟通过对高脂膳食肥胖大鼠进行长期规律的有氧运动,探讨运动干预对脂肪组织perilipin A mRNA表达的影响。
1 材料与方法
1.1 实验动物及饲料
健康清洁级3~4周龄离乳SD雄性大鼠60只,由华中科技大学同济医学院动物中心提供,体重60~95克。普通饲料由湖北省预防医学研究院提供。高脂饲料配方如下:50%标准饲料、15%猪油、10%白糖、5%奶粉、10%蛋黄、5%花生仁、5%芝麻油。
1.2 方法
1.2.1 肥胖与肥胖抵抗大鼠模型建立。SD大鼠适应性喂养3天后,随机分为2组,普通饲料喂养对照组(C)10只,高脂饲料诱导组50只,给予高脂饲料,自由进食、进水。大鼠肥胖判断标准:大鼠的体重超过普通饲料喂养大鼠平均体重的1.4倍标准差即可作为实验肥胖大鼠[3]。经高脂饲料喂养大鼠7周后,根据体重增加量,排名前16只大鼠拟定为肥胖大鼠。
1.2.2 肥胖与肥胖抵抗大鼠分组及干预。经过筛选出的肥胖大鼠随机分为两组(每组8只):肥胖不运动组(Ob)和肥胖运动组(Ob-E),第8周开始进行运动干预与喂养普通饲料,自由进食、进水。
有氧运动方案:大鼠专用电动跑台,坡度5°,速度15.2 m/min,相当于64±4.5%VO2max强度[4],并电击驱赶。运动时间60min,每周6次,周日休息,持续八周。
1.3 标本采集和保存
禁食12h后,将大鼠用戊巴比妥那腹腔麻醉,心脏取血后,分离血清,迅速取出肾周脂肪组织和睾周脂肪组织并称重,用铝箔纸包好,投入液氮中速冻,之后转入-80℃冰箱保存。
1.4 测定方法
1.4.1 血清生化指标的测定。总胆固醇(TC)、甘油三酯(TG)、高密度脂肪蛋白 (HDL-C)、低密度脂蛋白 (LDLC)由全自动生化分析仪测定,按试剂盒说明进行。
1.4.2 大鼠腹内脂肪组织perilipin A mRNA表达的测定。
1.4.2.1 主要试剂与仪器。Trizol Reagent购自美国Invitrogen公司,逆转录试剂盒购、实时荧光定量PCR试剂盒均购自日本TOYOBO公司。PCR仪(Life express Thermal Cycle)为杭州博日科技有限公司生产,荧光定量PCR仪(SLAN荧光定量PCR检测系统)为上海宏石医疗科技有限公司生产。
1.4.2.2 引物:由Invitrogen Biotechnology Co.,LTD中国公司合成,perilipin A上游引物:5′-GGAGACAGATGAGGAGGAAG-3′,下游引物:5′-GCTGGTGTGAGGTGTAGGAT-3′(NM_013094),产物207bp;β-actin上游引物:5’-CGTTGACATCCGTAAAGACCTC-3’,下游引物:5′ - TAGGAGCCAGGGCAGTAATCT - 3′ (NM031144),产物110bp。
1.4.2.3 PCR扩增。95℃预变性1min,95℃变性15s→58℃退火20s→72℃延伸20s,40次循环;72℃末段延伸5min;溶解曲线:72℃→95℃,每20s升温1℃。
1.4.2.4 结果处理。采用比较阈值法表示目的基因的相对表达,即ΔΔCt法:目的基因的相对表达量=2-△△Ct
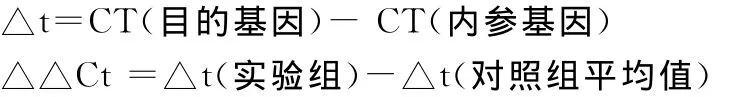
2-△△Ct表示的是实验组目的基因的表达相对于对照组的变化倍数。
1.5 统计学处理
统计分析使用13.0版SPSS统计软件完成。结果以“平均数±标准差”(±s)表示。统计分析采用单因素方差分析和独立样本t检验。P<0.05表示为显著性差异,P<0.01表示为非常显著性差异。
2 研究结果
2.1 运动对肥胖大鼠的体重、腹内脂肪、脂体比及血脂水平的影响
结果显示(见表1),与对照组(C组)相比,肥胖不运动组(Ob组)大鼠体重显著升高(392.88±21.86vs 462.00±31.90,P<0.05);腹内脂肪总量显著明显升高(9.56±0.70 vs 6.33±0.74,P<0.01);脂体比显著升高(2.05±0.23vs 1.61±0.13,P<0.05);血清 TG显著升高(0.49±0.10vs 0.38±0.09,P<0.05)。
8周跑台运动后,与Ob组相比,肥胖运动组(Ob-E组)大鼠腹内脂肪总量相比显著降低(7.44±1.06vs 9.56±0.70,P<0.05);脂体比相比显著降低(1.66±0.17vs 2.05±0.23,P<0.05)。血清TG相比 Ob组显著降低(0.41±0.05vs 0.49±0.10,P<0.05);Ob-E组血清 HDL-c显著高于C组(1.03±0.17vs 0.88±0.13,P<0.05)。
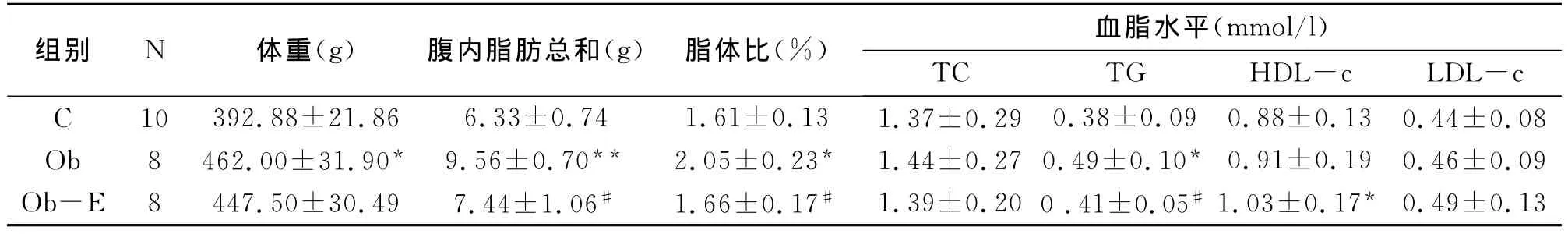
表1 各组大鼠检测指标比较表
2.2 运动对肥胖大鼠的脂肪组织perilipin A mRNA表达水平的影响
实验结果显示(见图1,图2),肥胖不运动组(Ob组)大鼠腹内脂肪组织perilipin A mRNA表达量是C组的1.96倍(1.96±0.89,P<0.05);肥胖运动组(Ob-E组)大鼠腹内脂肪组织perilipin A mRNA表达相比Ob组显著降低(1.11±0.79vs 1.96±0.89,P<0.05)见图1。
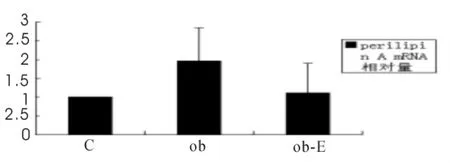
图1 大鼠腹内脂肪组织perilipin A mRNA表达示意图
图2 perilipin A扩增曲线示意图(1号样本)
3 分析讨论
3.1 运动对肥胖大鼠的体重、腹内脂肪、脂体比及血脂水平的影响分析
机体脂肪细胞过度积累、肥大,造成机体脂肪含量不断增加,形成肥胖,最终会导致机体的健康水平下降和生理机能的紊乱。肥胖与糖尿病、高血压、血脂异常、动脉硬化等多种疾病高度相关[5]。肥胖是由于过多的热能摄入与身体活动减少之间的不平衡造成的,田吉明等[6]对SD雄性大鼠进行10周高脂饮食可以引起大部分大鼠体重增加、脂肪重量和内脏重量增加,形成高脂饮食诱导肥胖大鼠。李世成等[7]对SD雄性幼鼠进行3周的高脂饲料喂饲,部分大鼠即达到肥胖标准。本研究测得肥胖大鼠,相较于普通饮食大鼠,肥胖不运动组(Ob组)大鼠体重、腹内脂肪总量、脂体比显著升高,提示通过高脂饮食的饲喂,Ob组大鼠有明显的内脏肥胖。
长期低强度有氧运动能够降低血清TC(总胆固醇)、TG(甘油三酯),使 HDL-c(高密度脂蛋白)升高,LDL-c(低密度脂蛋白)降低[8]。何玉秀等[9]研究发现9周有氧游泳运动后,雄性大鼠体重增加量减少,腹部脂肪垫(肾周脂肪垫和附睾脂肪垫)的脂肪积累减少。本研究通过8周的有氧运动跑台运动方案干预,测得大鼠Ob-E组大鼠腹内脂肪总量、脂体比、血清TG相比Ob组显著降低,HDL-c显著升高,说明8周有氧运动对改善内脏肥胖提高体质量、改善血脂水平有显著效果。
3.2 运动对高脂饮食肥胖大鼠脂肪组织perilipin基因表达的影响分析
Perilipin是特异表达于脂肪细胞和类固醇生成细胞脂滴表面的一种磷蛋白,作为一种“分子开关”,它可以直接参与脂质代谢的分子调控,胰岛素、儿茶酚胺、肾上腺素等可通过引起perilipin磷酸化而启动脂肪分解[10]。在脂质分解代谢中有双重调节作[11]:①在基础状态下,它被认为是脂质分解的限速酶即激素敏感性脂肪酶(HSL)通道的屏障,从而保护甘油三酯(TG)免遭分解;②在刺激状态下(如肾上腺素受体傲动剂诱导的脂解作用),Perilipin被cAMP依赖的蛋白激酶A磷酸化,使HSL移位,从而促进甘油三酯分解。
研究表明perilipin与肥胖症、2型糖尿病及动脉粥样硬化均密切相关。田吉明等[6]实验证明饮食导致大鼠肥胖与其脂肪组织HSL活性和蛋白表达下调以及perilipin蛋白表达上调,因而认为perilipin蛋白表达与促进脂肪储存有关。Tansey J T[12]等研究发现perilipin基因敲除小鼠出现了机体氧化、能量利用增加,而脂质合成减少,能够抵抗因高脂饮食所导致的肥胖。黄大祥[13]实验表明perilipm A mRNA在肥胖症和2型糖尿病患者皮下和网膜脂肪组织表达较正常人高。有研究数据表明,perilipin A基因具有抗动脉粥样硬化的作用,perilipin A基因敲除小鼠在没有血压及血脂水平升高的情况下,出现了动脉粥样硬化几率升高的现象[14]。本研究实验中Ob组大鼠腹内脂肪组织perilipin A mRNA表达显著高于对照组,也验证肥胖大鼠脂肪组织中perilipin A的基因表达上升。
关于运动训练对perilipin基因表达的影响的研究报道较少,并且报道结果不尽一致。田吉明[15]采用电动动物跑台运动,发现一次急性运动可改善肥胖大鼠运动后血糖、血脂,这种改变可能与其脂肪组织 HS L基因表达上调以及腹部脂肪组织perilipin基因表达基因表达下调有关。Petridou A等[16]研究报道经过8周的跑台运动后,运动组大鼠与未训练的大鼠相比,在肝脏有较低的脂肪酸合成酶,在附睾脂肪中有较高的脂肪酸合成酶、HSL和perilipin,在皮下脂肪有较高的HSL。两组大鼠的肝脏、肌肉或脂肪中PPAR浓度没有显著变化。本研究发现在进行8周的跑台运动方案干预后,Ob-E组大鼠腹内脂肪组织perilipin A mRNA表达相比Ob组显著降低。但也有与此不同研究结果,Chapados N等[17]关于运动训练对内脏脂肪和非内脏脂肪离体脂解的影响以及它们与周脂索浓度的关系的研究中发现,两组雌性大鼠均以高脂饲料喂养8周,一组无运动,另一组运动8周,而结果显示浓度没有受到运动的影响。因此,运动对perilipin A的影响还有待进一步进行研究。
4 结 论
通过本实验研究可知,高脂饮食致使大鼠腹部脂肪增加,血脂升高,腹部脂肪组织perilipin基因表达升高,而长期有氧运动可改善高脂饮食所致的肥胖大鼠的腹部脂肪减少,血脂水平改善,并且降低了腹部脂肪组织perilipin A基因表达。
[1]Hubert H,Feinleib M,McNamara P,Castelli WP:Obesity as an independent risk factor for cardio-vascular disease;a 26yearfollow-up of participants in the Framingham Heart Study.Circulation 1983,67:968-977.
[2]Londos C,Brasaemle DL,Schultz CJ et al.(1999)On the control of lipolysis in adipocytes.Ann NY Acad Sci 892:155-168.
[3]杨锡让,等.运动与身体成份——运动控制肥胖的机制研究[M].北京:北京体育大学出版社,1998:1-3.
[4]Bedford TG,Tipton CM,Wilson NC,et a1.Maximumoxygen consumption of rate and its changes with various experimental procedures[J].J Appl Physiol,1979,47(6):1278-1283.
[5]Bensimhon DR,Kraus WE,Donahue MP Obesity andphysical activity:a review.Am Heart[J]2006,151:598-603.
[6]田吉明,汪军,王瑞元.高脂饮食对大鼠脂肪组织脂解调节因素的影响[J].成都体育学院学报,2009,35(7):60-64.
[7]李世成,邵雄杰,等.运动对肥胖大鼠脂肪细胞脂联素mRNA表达的影响[J].西安体育学院学报2009,26(2):206-209.
[8]Mitsugi Motoyama,Yoshiyuki Sunami et al The effects of long-term low intensity aerobic training and detraining on serum lipid and lipoprotein concentrations in elderly men and women[J].Eur J App Physiol,1995,70:126-131.
[9]何玉秀,白文忠,姚玉霞,丛斌.运动对腹部脂肪积累及肥胖基因表达的影响[J].体育科学,1998,18(4):69-72.
[10]Dawn L.Brasaemle,Vidya Subramanian,Anne Garcia,et al Perilipin A and the control of triacylglycerol metabolism [J]Mol Cel Biochem 2009,326:15-21.
[11]Zhang HH,Souza SC,Muhro Kv,et a1.Lipase-selective functional domains of perilipin A differentially regulate constitutive and protein kinase A-stimulated lipolysis.J Biol Chem.2003,278:51535-51542.
[12]Tansey J T,Sztalryd C,Gruia-Gray J,et a1.Perilipin ablation results in a lean monse witIl aberrant adipocyte lipolysis,enhanced leptin production,and resistance to diet-induced obesity[J].J PNAS,2001,98(11):6494-6499.
[13]黄大祥.Perilipin在肥胖症和2型糖尿病患者脂肪组织中的表达[D].安徽医科大学,2006:29-30.
[14]Dominique Langlois Fabien Forcheron1et al Increased Atherosclerosis in Mice Deficient in Perilipin 1Lipids in Health and Disease 2011,10:169.
[15]田吉明,汪军,张瑞林.一次性运动对肥胖大鼠脂肪组织HSL和perilipin基因表达及血脂的影响[J].中国体育科技,2008,44(3):111-114.
[16]Petridou A,Tsalouhidou S,Tsalis G,et a1.Longterm exercise increases the DNA binding activity of peroxisome proliferator-activated receptor gamma in rat adipose tissue[J].Metabolism,2007,56(8):1029-1036.
[17]Chapados N,Collln P,Imbeauh P,et a1.Exercise training decreases in vitro sfimulated lipolysis in a visceral(mesenteric)but not in the retroperitoneal fat depot of high-fat-fed rats[J].Br J Nutr.2008,20:1-8.
