环境条件对转基因高粱Cry1Ab蛋白含量的影响
2012-07-31卢美贞崔海瑞
卢美贞,崔海瑞
(1.浙江工业大学 化学工程与材料学院,浙江杭州 310014;2.浙江大学原子核农业科学研究所,浙江杭州 310029)
苏云金芽孢杆菌(Bacillus thuringiensis,Bt)在芽孢形成时可产生具有杀虫活性的晶体蛋白[1],被称为杀虫晶体蛋白或 δ-内毒素[2]。Schnepf等[3]首次成功地克隆了第1个编码Bt杀虫晶体蛋白基因,揭开了利用基因工程培育抗虫转基因植物的序幕。目前它已成为植物抗虫基因工程中应用最广泛的抗虫基因。
浙江大学原子核农业科学研究所首次成功地将来自Bt密码子优化的以玉米泛素为启动子的Cry1Ab抗虫基因转入高粱中[4],并经农业部批准进入中间试验阶段。转基因高粱的获得为这一作物的品种改良提供了抗虫资源。为有效和合理地利用这一资源,有必要了解不同光照时间、不同温度和不同水平氮肥处理下,Cry1Ab基因在高粱中的表达情况。本研究以转Cry1Ab基因高粱SR16-2和SR24-2为材料,研究不同光照时间、不同温度和不同水平氮肥处理下,Cry1Ab基因在高粱中的表达特性。
1 材料与方法
1.1 材料
研究所用的材料为高粱原受体亲本115R及其转入Cry1Ab基因的2个稳定转化株系,这2个株系处于 R5代,编号分别为 SR16-2和 SR24-2,它们分别来自2个独立转化株,系采用农杆菌介导法培育而成[4]。
1.2 处理设计
1.2.1 光照处理
取SR16-2和SR24-2株系干种子各100粒,进行常规浸种、催芽,种子萌动后放在人工培养室28℃恒温下培养,设3次重复,每重复20粒种子,设置的光照时间分别为 10,12,14,16和18 h,待幼苗长到3叶期时,各处理随机取幼苗(去根)5个测Cry1Ab蛋白含量。
1.2.2 温度处理
取SR16-2和SR24-2株系干种子各100粒,进行常规浸种、催芽,种子萌动后放在光照培养箱中培养,每日相同光强光照12 h,设计了5个温度水平,分别为20,25,30,35和40℃,湿度在75%左右,设3次重复,每重复20粒种子,待幼苗长到3叶期时,各处理随机取幼苗(去根)5个,测Cry1Ab蛋白含量。
1.2.3 施氮水平处理
取SR16-2和SR24-2株系干种子各100粒,进行常规浸种、催芽,萌动后将种子播种于网室,分别设置 4个基肥纯氮水平,0,300,450和600 kg·hm-2,进行常规栽培,在拔节期各处理分别随机选5株取叶片测Cry1Ab蛋白含量。
1.3 Cry1Ab蛋白测定
以上所取样品洗净后称重,在液氮中磨成粉状,放入离心管中,按4 μL·mg-1的比例加适量蛋白提取缓冲液(试剂盒随带),充分混匀后置于4℃浸提3 h,5 000 r·min-1离心 5 min,上清液或其稀释液即可用于测定。将100 μL样品提取液加入到已包埋好抗体的96孔板中,同时加入不同浓度(0,0.5,2.5,5.0 ng·mL-1)的 Cry1Ab蛋白标准溶液,按 Cry1Ab/Cry1Ac板式试剂盒(美国ENVIROLOGIX公司)提供的方法进行测定,最后根据酶标仪(SpactronMax190,德国)在波长为450 nm下D的读数,参照标准曲线和提取液稀释倍数计算出各器官组织Cry1Ab杀虫蛋白含量,以样品中所含的Cry1Ab蛋白含量来表示(ng·g-1),测定时各设3次重复。
2 结果与分析
2.1 不同光照时间对Cry1Ab蛋白量的影响
对不同光照长度处理的Cry1Ab蛋白含量测定结果如图1。
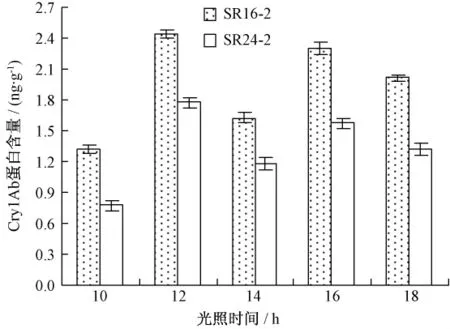
图1 不同光照时间下转基因高粱幼苗中Cry1Ab蛋白含量
由图1可见,在10,12,14,16和18 h等不同光照时间处理后,2个转基因株系的Cry1Ab蛋白含量平均分别为 1.05,2.10,1.40,1.94和1.67 ng·g-1,以光照12 h时的 Cry1Ab蛋白含量最高,光照10 h的表达量最低,二者相差1倍以上,表现趋势为12 h>16 h>18 h>14 h>10 h,且2个转基因株系表现出相同的趋势。说明光照长度对Cry1Ab蛋白表达量具有一定的影响。
2.2 不同温度处理对Cry1Ab蛋白量的影响
对Cry1Ab蛋白含量测定结果显示,不同温度处理对 Cry1Ab蛋白含量也有较大影响(图2)。25℃培养时叶片中 Cry1Ab蛋白含量最高,达到2.24 ng·g-1,温度升高,Cry1Ab蛋白含量呈现下降趋势,40℃最低,只有0.44 ng·g-1,20℃时叶片中Cry1Ab蛋白含量比35℃略高。可见温度也是影响Cry1Ab蛋白表达水平的一个重要环境因素。
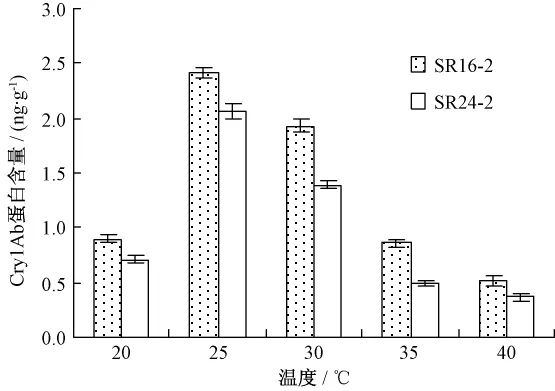
图2 不同温度处理下转基因高粱幼苗中Cry1Ab蛋白含量
2.3 不同施肥水平对Cry1Ab蛋白量的影响
图3是对不同施氮水平下Cry1Ab蛋白量的测定结果。当以 300,450,600 kg·hm-2不同施氮水平作基肥处理时,SR16-2转基因高粱株系叶片中的Cry1Ab蛋白含量比未施基肥的对照分别显著提高85.40%、48.66%和12.90%;在另一转基因高粱株系SR24-2中,施肥后的Cry1Ab蛋白含量与对照相比差异更大,分别提高了200.42%,50.42%和39.58%。结果表明,施肥可以促进Cry1Ab蛋白的合成,作基肥的以300 kg·hm-2纯氮效果最佳。
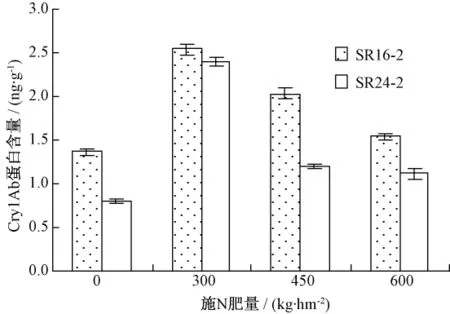
图3 不同施N基肥量下转基因高粱Cry1Ab蛋白含量
3 小结与讨论
Bt蛋白含量的高低直接影响着转 Bt基因植物的抗虫效果,而环境因素对Bt基因的表达起着重要的调节作用[5],尤其是当 Bt基因处于诱导型启动子控制下时,这种影响更为突出。因此,阐明环境因素对Bt基因表达水平的影响对转Bt基因植物的合理栽培与利用具有重要的指导作用。本研究的结果表明,光照、温度和氮肥对Cry1Ab蛋白表达量都具有明显的影响,为转Cry1Ab基因高粱的进一步应用提供了理论依据。
温度对外源基因的表达起着重要的调节作用[6]。Takimot等[7]曾报道玉米泛素启动子的表达不受系统调节,而是受独立的对热击或物理胁迫产生反应的细胞调节,在受胁迫情况下,该启动子会改变它的表达特性。此外,Meyer[8]也曾报道过温度会影响转基因矮牵牛中35S启动子的甲基化。本研究所用转基因高粱中的Cry1Ab基因是由玉米泛素启动子驱动的,研究结果显示,温度对转基因高粱Cry1Ab蛋白含量有较大的影响,高温不利于Cry1Ab蛋白的表达,这与吴刚[9]对玉米泛素启动子控制的Cryab在转基因水稻中表达量受温度调节的研究结果相一致,都证明了 Takimoto等[7]的结论,也与夏兰芹等[10]报道棉花中35S启动子引导的Bt基因所表达的杀虫蛋白量在高温胁迫后急剧下降的结果相似。此外,在转Bt基因棉花中也有因气温反常导致抗虫性下降的报道[11]。因此,在转Bt基因抗虫植物的种植过程中,应根据温度预报情况加强监测,在高温期到来之际综合采用其他防治措施。
合理施肥是作物栽培管理的关键。施肥直接促进根系对氮素等养分的吸收,进而影响植株体内蛋白质的合成。在转Bt基因棉花中,王家宝等[12]报道追肥可提高叶片中的Bt蛋白含量,且随追肥量的增加而增加;周冬生等[13]也报道随追肥量的增加转Bt基因棉抗虫性增强。本研究结果表明,施氮基肥有助于Cry1Ab蛋白含量的提高,但是随着施氮量的增加Cry1Ab蛋白含量并未呈现增长的趋势,反而有所下降。由此可见,作基肥与追肥的效果不同。因此,对转Bt基因抗虫植物的施肥,不但要控制氮肥的量,还应讲究施肥方式。
光照直接影响着植物的生长、发育。有关光照对转基因表达的影响,在转玉米A1基因的矮牵牛中已有报道[7],但光照是否影响 Bt基因在转基因植物中的表达水平尚未见报道。高粱属短日照植物,临界光周期约为12 h。本研究中发现光照12 h时转基因高粱叶片中的Cry1Ab蛋白量最高,光照长于或短于这一时间均导致Cry1Ab蛋白含量的降低。因此,要获得较好的抗虫效果,在种植转Bt基因高粱时应从光照长度这一角度考虑其合适的地域。
[1]Hannay C L.Crystalline inclusions in aerobic spore-forming bacteria[J].Nature,1953,172:1004.
[2]Hannay C L.The protein crystals of Berliner[J].Can J Microbiol,1955,1:694 -710.
[3]Schnepf H E,Whiteley H R.Cloning and expression of the Bacillus thuringiensis crystal protein gene in Escherichia coli[J].Proc Natl Acad Sci USA,1981,78:2893-2897.
[4]张明洲.农杆菌介导高梁转化体系的建立与 Bt(Bacillus thuringiensis)抗虫基因的导入[D].杭州:浙江大学,2002.
[5]Sachs E S,Benedict J H.Pyramiding Cry1A(b)insecticidal protein and terpenoids in cotton to resist tobacco budworm[J].Environmental Entomology,1998,25(6):1257-1266.
[6]周桂生,周福才,谢义明,等.温度胁迫对转 Bt基因抗虫棉毒蛋白的表达和棉铃虫死亡率的影响[J].棉花学报,2009,21(4):302-306.
[7]Takimoto I,Christensen A H,Quail P H,et al.Non-system expression of a stress-responsive maize polyubiquitin gene(Ubi-1)in transgenic rice plants[J].Plant Molecular Biology,1994,26:1007-1012.
[8]Meyer P.Endogenous and environmental factors influence 35S promoter methylation of a maize Al gene construct in transgenic petunia and in color phenotype[J].Molecular and General Genetic,1994,6:1410-1414.
[9]吴刚.cry1Ab基因在转基因水稻中的遗传、表达与沉默[D].杭州:浙江大学,2000.
[10]夏兰芹,郭三堆.高温对转基因抗虫棉中 Bt杀虫基因表达的影响[J].中国农业科学,2004,37(11):1733-1737.
[11]Kaiser J.Pests over whelm Bt cotton crop[J].Science,1996,223:423-427.
[12]王家宝,王留明,沈法富,等.环境因素对转 Bt基因棉Bt杀虫蛋白表达量的影响[J].山东农业科学,2000,(6):4-6.
[13]周冬生,吴振廷,王学林等.施肥量和环境温度对转 Bt基因棉抗虫性的影响[J].安徽农业大学学报,2000,27(4):352-357.
