盐酸法舒地尔对脑缺血再灌注损伤大鼠神经细胞凋亡和细胞色素C释放的影响
2012-07-20庄育刚陈艳清张翔宇
陈 胜,彭 沪,庄育刚,陈艳清,张翔宇
(同济大学附属第十人民医院急诊危重病科,上海 200072)
基础研究表明,细胞凋亡与脑缺血再灌注损伤有着密切的关系,凋亡是脑缺血再灌注损伤后神经元死亡的重要方式之一。最近研究发现,盐酸法舒地尔是一种新型Rho-激酶抑制剂(RKI),可以有效预防和缓解脑血管痉挛,具有强大的扩血管作用和保护缺血脑组织的作用,但其是否具有减轻脑神经细胞凋亡的作用还未完全清楚。本实验旨在通过观察法舒地尔对大鼠急性脑缺血再灌注损伤后大鼠海马CA1区神经细胞病理形态和线粒体超微结构改变,以及对细胞色素C(cytC)介导的线粒体凋亡通路中的关键蛋白cytC表达的影响,从细胞凋亡及其信号转导通路角度探讨法舒地尔对脑缺血再灌注损伤的保护作用。
1 材料与方法
1.1 实验动物与分组
雄性健康SD大鼠72只,体质量250~300 g,由同济大学医学院实验动物中心提供。随机分成3组:假手术组(S组,n=32);缺血再灌注组(IR组,n=32);盐酸法舒地尔组(F组,n=24)。三组按再灌注时间再分为3个亚组,即缺血10 min后分别再灌注3、12、24 h,每个亚组动物均为8只。实验前3 d适应实验室环境,大鼠自由饮水和进食。
1.2 试剂
盐酸法舒地尔注射液(规格2 ml/30 mg),由天津红日药业股份有限公司提供,cytC免疫组化试剂盒购自武汉博德生物公司。
1.3 模型制作
采用改良的四血管阻滞法建立大鼠全脑缺血再灌注模型 。①称体质量,麻醉:10%水合氯醛0.35 ml/100 g腹腔注射麻醉待其四肢瘫软后取出。②分离或损毁椎动脉:将大鼠腹卧位固定于立体定位仪上,鼠头与躯干向前屈曲30°,颈后正中切口长约1 cm,暴露第一颈椎两侧横突翼小孔(椎动脉在入脑前从此孔下通过)。用电凝器凝闭双侧椎动脉,若仍有较多血液渗出,用小棉球压堵翼小孔1~2 min即可止血。消毒后缝合肌肉及皮肤,回笼。③分离或夹闭双侧颈总动脉及脑电监测:24 h后称重,10%水合氯醛0.35 ml/100 g腹腔麻醉,待大鼠进入麻醉状态后将银制电极分别插入额部,鼻颞部及背部皮下供脑电监测。描记正常脑电图记录打标并保存。继之大鼠重新背位固定,颈正中切口2~3 cm,颈动脉三角处牵开胸锁乳突肌,分离、暴露两侧颈总动脉,在颈总动脉下留置一根5-0丝线待用,缺血时提拉两端线头,待动物清醒后以动脉夹夹闭双侧颈总动脉,造成全脑缺血,并及时瞄记脑电图并记录时间,脑电图波幅下降至缺血前波幅25%以下或呈一条直线为全脑模型成功标志。10 min后松开动脉夹恢复脑血流即为再灌注。约15 min后脑电图逐渐回复至原图形波幅高度,缝合颈部创口,回笼饲养。④分组造模:S组仅暴露双侧椎动脉不灼闭,分离出双侧颈总动脉而不夹闭。F组在再灌注前立即腹腔注射盐酸法舒地尔(30 mg/kg)。S组,I/R组给予等量生理盐水腹腔注射。⑤在全脑缺血实验过程中,用白炽灯照射动物以维持肛温37℃左右直到恢复活动。再灌注前后观察动物脑电图,翻正反射及瞳孔变化等以判断是否发生全脑缺血再灌注损伤。
1.4 模型成功标准
手术操作中无大出血,双侧颈总动脉夹闭10 min内翻正反射消失,吊尾圆锥运动无挣扎,瞳孔散大,提示全脑缺血。脑电图波幅下降至缺血前波幅25%以下或一直线提示形成全脑缺血再灌注损伤,缺血10 min松开血管夹血流恢复1~2 min后上述症状消失,约15 min后脑电图逐渐回复至原图形波幅高度;存活期内未见癫痫发作表明模型制作成功,否则模型失败,弃去不用。
1.5 标本采集
各亚组动物分别于再灌注3、12、24 h后腹腔注射10%水合氯醛0.35 ml/100 g麻醉,开胸暴露心脏,以0.9%生理盐水200 ml经心脏快速灌注冲洗,随后以体积分数为4%的多聚甲醛磷酸盐缓冲液(PBS,40 ℃,pH7.14)200 ml经心脏灌注约30 min。立即冰上取脑,在脑冠状切面,视交叉后约1.5 mm切开脑组织,向后切取5 mm标本。4℃ 4%多聚甲醛固定24 h,常规冲洗、脱水、透明、包埋,连续冠状切片,片厚5 μm,做免疫组化切片。其中,每组随机抽取再灌注24 h亚组大鼠两只,过度麻醉后取视交叉后1.5 mm脑海马组织厚浸泡于4℃ 3%戊二醛中固定脱水切片包埋染色做透射电镜观察。
1.6 指标检测
1.6.1 cytC蛋白检测 采用免疫组化SP法检测cytC蛋白的表达,严格按试剂盒说明进行操作。DAB显色,显微镜下控制反应时间,常规封片。细胞浆中出现棕黄色颗粒、胞核不着色者为阳性细胞。采用HPIAS-2000图像分析系统(同济大学医学院病理教研室提供)对切片进行图像分析,分析过程中所有切片放大400倍,光源强度相同,测试每张切片cytC阳性细胞的平均光密度值。
1.6.2 光镜观察 苏木素伊红(H-E)染色,光镜下观察每组再灌注24 h亚组海马CA1区的神经细胞形态学改变。
1.6.3 电镜观察 戊二醛和锇酸双重固定,脱水、环氧树脂包埋,行超薄切片,采用醋酸铀和硝酸铅双重染色,日立H-600型透视电镜(同济大学医学院病理教研室提供)观察大鼠海马CA1区线粒体超微结构变化。
1.7 统计学分析
采用SPSS13.0统计软件进行数据处理。所有数据用±s表示,组内和组间比较采用单因素方差分析(Post Hoc Tests,两两比较采用SNK法),并定义P<0.05差异有统计学意义,P<0.01差异有显著的统计学意义。
2 结 果
2.1 cytC免疫组化结果
①大鼠脑缺血再灌注后海马CA1区cytC在再灌注3 h即有明显表达,于再灌注12 h达高峰,以后表达逐渐降低;②S组海马CA1区仅有极少量的cytC阳性细胞,各亚组间比较无明显差异(P>0.05),与S组比较,IR组和F组各时间点海马CA1区的cytC蛋白表达均明显升高,差异具有显著的统计学意义(P<0.01);③与IR组比较,F组各时间点海马CA1区的cytC蛋白表达均明显降低,差异具有统计学意义(P<0.05或P<0.01)(图1、2)。
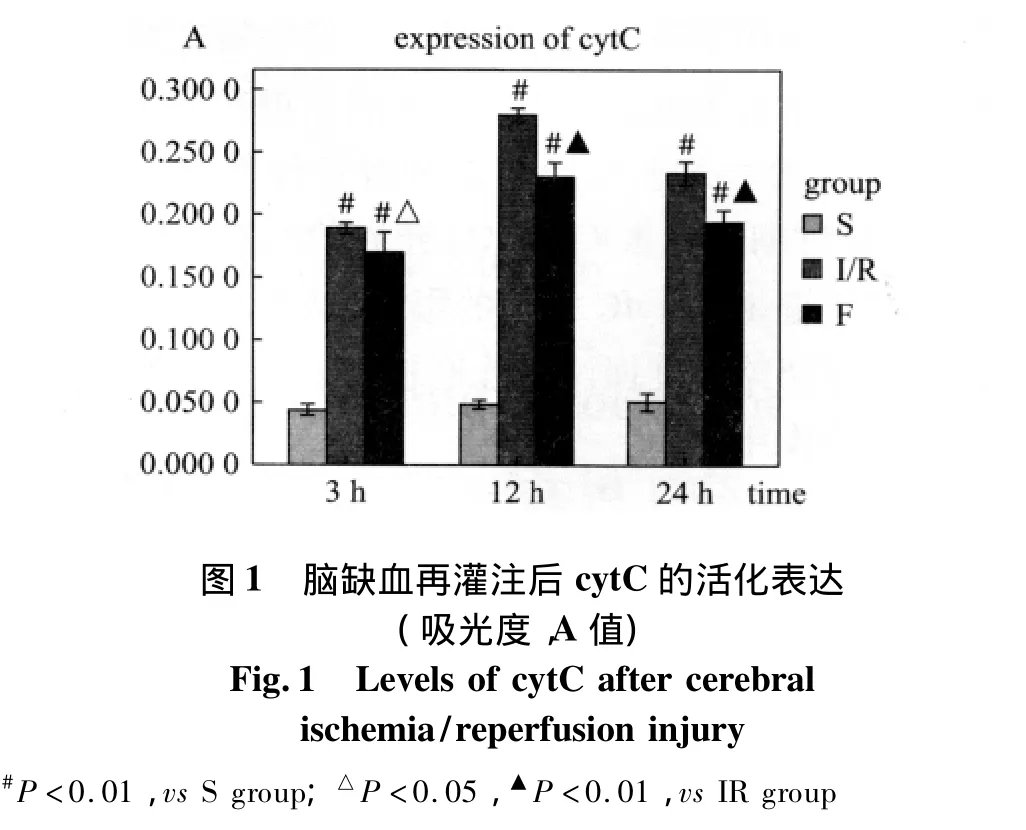
2.2 H-E染色光镜观察
S组海马CA1区神经细胞形态基本正常,细胞无肿胀,排列整齐,数目无明显变化(图3A)。与S组比较,IR组神经细胞受损严重,细胞水肿,排列混乱,正常神经元数目明显减少,可见大量炎症细胞浸润,神经基质明显肿胀,细胞间隙明显增宽。部分细胞胞核浓缩深染,胞浆强嗜酸性变,核固缩明显,可见核仁消失,核碎裂,核溶解(图3B)。而F组较IR组神经元受损情况明显减轻,神经细胞呈轻度皱缩或肿胀,排列较整齐,细胞间隙略增宽,炎症细胞浸润也较IR组明显减少(图3C)。
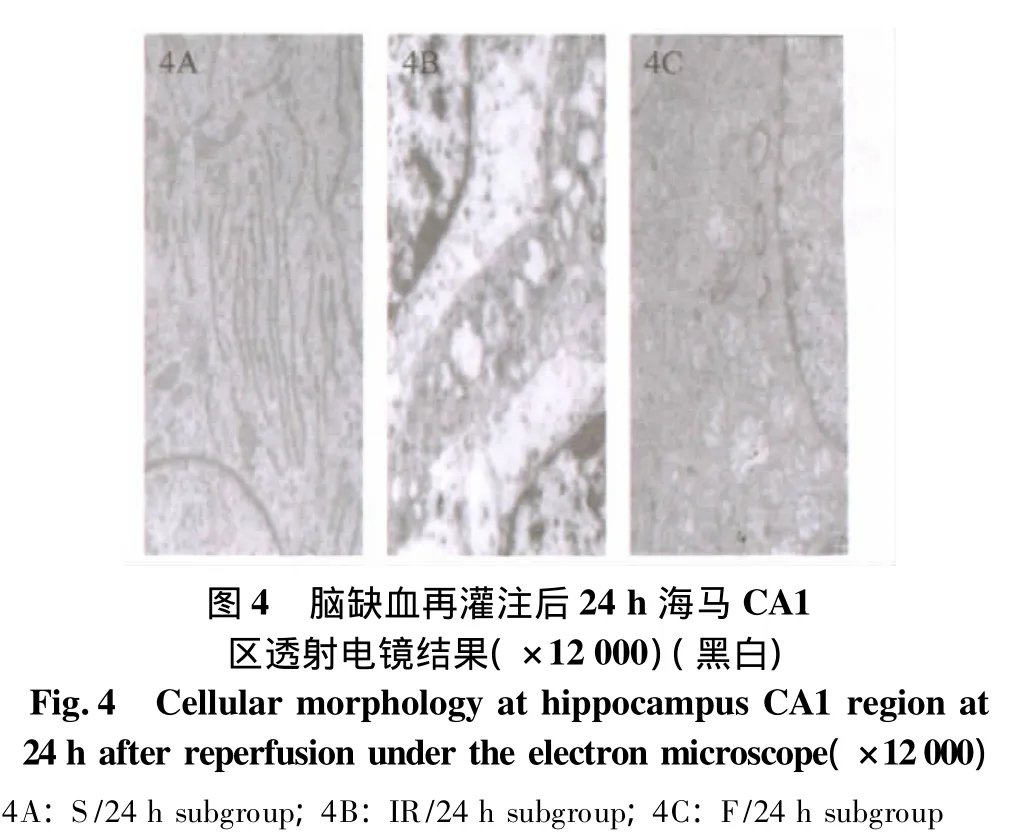
2.3 透射电镜观察
S组海马CA1区的超微结构大致正常:核膜光滑,核仁清晰,电子密度均匀,神经细胞核呈卵圆形,无切迹,胞质内有大量线粒体,切面为椭圆形,内有基质与排列密杂的嵴(图4A)。IR组:神经细胞损害严重,细胞质溶解破坏明显,染色质凝集,核仁边移,核膜迂曲、凹陷显著,连续性中断。胞质中线粒体多数肿胀,呈烧瓶样或空泡样,其内部嵴断裂、脱失,基质颗粒脱落(图4B)。F组:细胞有一定程度肿胀、破坏,但较IR组明显减轻,核膜较平滑、连续性较好,电子密度较均匀,细胞核内可见染色质成点状凝集,核仁较清晰,线粒体形态基本规则,但仍有一定程度的肿胀,大多数嵴完整,排列较紧密,基质颗粒极少量脱落(见图4C)。
3 讨 论
在脑缺血再灌注损伤过程中,由caspase依赖的线粒体凋亡通路已经得到实验研究的证实[1]。其中,cytC和caspase-3起着关键的作用。cytC是第一种被发现的线粒体释放的促凋亡蛋白,在线粒体损伤后,cytC作为应激传感器被释放到胞质中,从而引发细胞执行凋亡程序[2]。近年大量基础研究表明,细胞凋亡是脑缺血再灌注损伤后神经元死亡的重要方式之一[3-4]。在脑缺血再灌注损伤过程中,由CytC介导的线粒体凋亡通路已得到实验研究的证实,CytC在其中起着关键的作用[5-6]。王晓东教授最先提出cytC在细胞凋亡中的作用[7]:他研究发现细胞受促凋亡因素刺激后,cytC迅速从线粒体内释放到胞浆中,启动细胞凋亡,cytC的释放与蛋白酶原的激活形成一个负反馈的自我放大回路。Chihoko等应用大鼠MCAO模型,发现胞浆中CytC 3 h 开始出现明显升高[8]。Taku Sugawara 等[9]研究表明,全脑缺血再灌注后2 h海马CA1区线粒体CytC释放入胞浆,并于12 h达高峰。同时,Zhao等[10]研究发现,全脑缺血再灌注后2 h海马CA1区线粒体CytC释放入胞浆,并于12 h达高峰。张宇红等[11]也发现脑缺血再灌注损伤后海马 CA1区CytC的表达明显增加。
本研究结果显示,大鼠脑缺血再灌注后海马CA1区cytC蛋白在再灌注3 h即有明显表达,于再灌注12 h达高峰,以后表达逐渐降低。脑缺血再灌注组CytC蛋白表达较假手术组显著增加,神经细胞病理形态和线粒体超微结构受损明显,提示Cyt介导的线粒体凋亡通路参与了脑缺血再灌注后神经元凋亡的调控。我们给予盐酸法舒地尔干预后,法舒地尔通过阻止部分线粒体内膜间隙cytC蛋白的释放和激活,稳定线粒体膜,减少线粒体内部嵴断裂和基质脱颗粒,保护线粒体的形态功能,从而减少细胞凋亡的发生,在线粒体水平上发挥脑保护作用。
盐酸法舒地尔,一种新型细胞内钙离子拮抗剂,具有强大的扩血管作用,很好地保护缺血脑组织。它作为细胞内钙离子拮抗剂,加速钙离子浓度的恢复,减少线粒体内钙积聚,维持线粒体的正常功能,阻止部分线粒体内膜间隙cytC蛋白的释放和激活,保持了线粒体细胞氧化代谢,从而减少乳酸的产生和促进乳酸的氧化,纠正酸中毒。细胞内钙超载在脑缺血再灌注损伤后神经细胞凋亡的发生发展中起到关键作用。脑缺血再灌注损伤中Ca2+超载是多种因素综合作用的结果,也是脑缺血再灌注损伤中各种因素作用的共同通路。细胞内Ca2+的升高参与了凋亡早期的信号转导和凋亡过程的执行阶段。细胞凋亡早期线粒体外膜通透性增加、Ca2+的摄入增加、跨膜电位降低、CytC和凋亡诱导因子Apaf-1和procaspase-9的释放等,最终导致Cyt介导的线粒体凋亡通路的发生[10]。
综上所述,盐酸法舒地尔对CytC介导的线粒体凋亡通路有干预作用,通过保护线粒体的形态功能,稳定线粒体膜,抑制cytC的释放和激活,从而减少细胞凋亡的发生,发挥脑保护作用,这可能是其改善缺血再灌注损伤的部分作用机制。关于盐酸法舒地尔治疗脑缺血再灌注损伤是否还存在其他机制以及有无剂量依赖性有待下一步的研究证实。
[1]Graham SH,Chen J.Programmed cell death in cerebral ischemia[J].J Cereb Blood Flow Metab,2001,21(2):99-109.
[2]Zemlyak I,Brooke SM,Singh MH,et al.Effects of overexpression ofantioxidants on the release of cytochrome c and apoptosis-inducingfactor in the model of ischemia[J].Neurosci Lett.2009,453(3):182-185.
[3]Dang S,Liu X,Fu P,Gong W,et al.Neuroprotection by local intra-arterial infusion of erythropoietin after focal cerebral ischemia in rats[J].Neurol Res,2011,33(5):520-528.
[4]Lznerowicz A, Chudoba P, Kamińska D,etal.Duration of brain death and cold ischemia time,but not warm ischemia time,increases expression of genes associated with apoptosis in transplanted kidneys from deceased donors[J].Transplant Proc,2011,43(8):2887-2890.
[5]Martelli AM. Virusesand apopotosis[J]. Cell Biochem,2001,82:634.
[6]Ding ZM,Wu B,Zhang WQ et al.Neuroprotective Effects of Ischemic Preconditioning and Postconditioning on Global Brain Ischemia in Rats through the Same Effect on Inhibition of Apoptosis[J].Int J Mol Sci,2012;13(5):6089-6101.
[7]萨其拉,李文彬,孙勇如,等.线粒体与细胞凋亡机制[J].国外医学分子生物学分册,2000,22(5):277-281.
[8]Liang BT.Protein kinase C-mediated preconditioning of cardiac myocytes:role of adenosine receptor and KATP channel[J].Am J Physiol,1997,273(2):847 - 853.
[9]Lindholm D,Eriksson O,Korhonen L.Mitochondrial proteinsin neuronaldegeneration[J]. Biochem Biophys Res Commun,2004,321(4):753 -758.
[10]Groenendyk J,Lynch J,Michalak M.Calreticulin,Ca2+,and calcineurin-signaling from the endoplasmic reticulum[J].Mol Cells,2004,17(3):383 -389.
[11]张宇红,汪锡金,叶民,等.黄连素抑制促炎症因子生成保护多巴胺能神经元[J].同济大学学报:医学版,2009,30(3):42 -45,64.
