A common variant in the precursor miR-146a sequence does not predispose to cholangiocarcinoma in a large European cohort
2012-07-07FlorentinaMihalacheAksanablingerMonicaAcalovschiTilmanSauerbruchFrankLammertandVincentZimmer
Florentina Mihalache, Aksana Höblinger, Monica Acalovschi, Tilman Sauerbruch, Frank Lammert and Vincent Zimmer
Homburg, Germany
Original Article / Biliary
A common variant in the precursor miR-146a sequence does not predispose to cholangiocarcinoma in a large European cohort
Florentina Mihalache, Aksana Höblinger, Monica Acalovschi, Tilman Sauerbruch, Frank Lammert and Vincent Zimmer
Homburg, Germany
BACKGROUND:Micro-RNAs (miRNAs) are small, non-coding RNA species considered to fine-tune basic cellular functions by modulating target gene translation and/or mRNA stability. A common G/C polymorphism (rs2910164) in the precursor (pre-)miR-146agene engaged in NF-κB signaling and apoptosis pathways has been reported to modulate the genetic risk of hepatocellular carcinoma by increased G-allelic production of maturemiR-146a. We investigatedrs2910164in a large Europeanbased cholangiocarcinoma (CCA) cohort.
METHODS:We recruited 182 CCA patients and 350 controls in three academic medical centers. Genotyping forrs2910164was performed by PCR-based assays with 5'-nuclease and fluorescence detection. Genotype frequencies were tested for consistency with the Hardy-Weinberg equilibrium using an exact test; allelic and genotypic differences between the patients and controls were assessed by the Chi-square test and Armitage's trend test. Exploratory subgroup analyses included gender, tumor localization (extra- versus intrahepatic CCA) and early-onset CCA.
RESULTS:Genotype distributions were consistent with the Hardy-Weinberg equilibrium. No significant differences in either allele or genotype distributions were detected between the CCA and control groups or the respective subgroups investigated. However, there was a trend for a protective effect of the heterozygous single-nucleotide polymorphism state GC, as indicated by an underrepresentation in the CCA group in general (29% vs 35%;P=0.18) and, in particular, for extrahepatic tumor sites (26% vs 35%; OR=0.67; 95% CI, 0.43-1.02;P=0.065).
CONCLUSIONS:Our data do not support a prominent contribution of the pre-miR-146asequence variant in the genetic predisposition to CCA. However, current studies functionally characterizingrs2910164have proposed that distinct repertoires of target genes are addressed by genotype-specific maturemiR-146aspecies. Given the detected trend towards a potentially protective role of GC heterozygosity, a subtle modulation of genetic CCA risk by the pre-miR-146aGC genotype may exist and should be evaluated further.
(Hepatobiliary Pancreat Dis Int 2012;11:412-417)
miR-146a; miR-SNP; biliary tract cancer; cholangiocarcinoma; genetic risk
Introduction
Micro-RNAs (miRNAs) are small non-coding RNA species (18-22 nucleotides in length) involved in fine-tuning basic cellular functions by modulating target gene translation and/or messenger RNA (mRNA) stability through sequence-specific binding to mRNA.[1]In general, single-nucleotide polymorphism (SNP) variation in miRNA genes has been considered uncommon owing to high evolutionary conservation across species. However, recently several functional variants in primary (pri-) or precursor (pre-)miRNAsequences, affecting their complex biogenesis, or within the target binding sites of the mature miRNA sequence, have been identified and are known as miRSNPs.[2]The functional G/C SNPrs2910164in the seed sequence of pre-miRNA-146aresults in mispairing of hairpin structures, thus causing aberrant processing and potential changes in the expression level of the mature miRNA sequence.[3]Interestingly, this variant, affecting pathways relevant to hepatobiliarycarcinogenesis such as nuclear factor-κB (NF-κB) signaling and apoptosis regulation, has been linked to the genetic risk of hepatocellular carcinoma in men via increased G-allelic production of maturemiR-146a.[4]However, in contrast, current data suggest that heterozygosity rather than homozygosity forrs2910164modulates the genetic susceptibility to papillary thyroid cancer.[3]In subsequent work, these authors have elegantly shown that GC heterozygosity yields three distinct species of maturemiR-146a, each of which has a specific repertoire of target genes, thereby offering an attractive explanation for the observed differences in phenotypes.[5]As a matter of fact, the functional annotation of thers2910164GC genotype lends credence to the genetic risk attribution to allele heterozygosity rather than homozygosity, representing a highly uncommon phenomenon in genetics, referred to as "overdominance".[6]Taken together, there are conflicting data on whether GG homozygosity or GC heterozygosity confers genetic risk of different cancer types.
Cholangiocarcinoma (CCA) is the second most common primary liver cancer, and malignant transformation of biliary epithelial cells is considered to originate from a multistep process in which chronic inflammation and cholestasis play critical roles.[7]Among others, major clinical risk factors include primary sclerosing cholangitis, viral hepatitis, metabolic syndrome and liver-fluke infections in Asian countries.[7,8]In addition to increased age as a common risk factor, there is a slight gender disparity in CCA epidemiology, with men showing 1.2-1.5-fold higher CCA incidence rates in comparison to women.[9]Genetic susceptibility factors affecting an individual's risk of developing malignant biliary tract disease are not well defined,[10]not least owing to the overall low CCA incidence, which complicates the establishment of informative study cohorts. However, it is assumed that the recognition of genetic factors relevant to cholangiocarcinogenesis may help identify novel carcinogenic pathways and individuals at high risk of disease potentially amenable to strategies aiming at early diagnosis. In this line, we demonstrated in our previous study that genetic CCA risk, in particular for extrahepatic CCA, is increased in carriers of thealpha1-antitrypsinZ allele.[11]Of note, aberrant expression of different sets of miRNA species has been reported in CCA; however, only few common genetic variants in evolutionarily conserved miRNA loci have been reported.[12,13]
Thus, our aim in the current study was to evaluate the putative role ofrs2910164in the seed sequence of pre-miR-146ain the genetic susceptibility to CCA.
Methods
Study design and population
We included 182 Caucasian CCA patients and 350 controls from Germany (University Hospitals of Bonn and Homburg: 152/290) and Romania (Cluj-Napoca: 30/60) in our study. Most cancer patients were recruited prospectively (n=128) whereas another 54 were enrolled retrospectively. Additional details of patient recruitment and blood sample acquisition are given in our previous paper.[11]Cancers were classified as either extra- or intrahepatic CCAs as defined by the British Society of Gastroenterology.[14]There was an unequivocal tissue diagnosis of CCA either by histology or cytology in 84% of participants, which was supported by adequate exclusion of extrabiliary cancers. In the remaining 16% of case subjects, CCA was diagnosed clinically after evaluation of cross-sectional and appropriate biliary imaging, follow-up data, and lack of potential alternative diagnosis such as postoperative strictures. The control group comprised individuals asked to participate in after negative colorectal cancer screening by colonoscopy (290 individuals) and/or negative abdominal ultrasound for gallstone disease (60 individuals). Written informed consent was obtained from all participants, and the local ethic committees approved the study, which was in accordance with the revisedDeclaration of Helsinki.
Genotyping procedure
We applied the QIAamp protocol (Qiagen, Hilden, Germany) for extraction of genomic DNA, which was calibrated to 5-50 ng/µL, using a NanoDrop ND-1000 spectrophotometer (NanoDrop Technologies, DE, USA). Solution-phase hybridization reactions with 5'-nuclease and fluorescence detection (TaqMan assays, Applied Biosystems, Foster City, CA, USA) were used for genotypingrs2910164on an ABI 7500 Fast Real-Time polymerase chain reaction (PCR) platform (Applera, Norwalk, CT, USA). Technical details were reported previously.[11]For validation of genotyping results, >10% of samples were genotyped in duplicate.
Statistical analysis
Data were expressed as mean and standard deviation (SD). Age and gender distributions in cases and controls were compared using Student'sttest.Pvalues <0.05 were considered significant. Compliance of genotype distributions in controls with the Hardy-Weinberg equilibrium (HWE) was assessed by an exact test (http://ihg.gsf.de/snps.html). The Chi-square test and Armitage's trend test were applied to compare allele and genotype frequencies between patients and controls incontingency tables. We used the SPSS software package (version 16.0: SPSS, Munich, Germany) for statistical analyses. Moreover, power calculations to assess for an adequate sample size were performed using the PS Power and Sample Size Calculation v.3.0 software package (http:// biostat.mc.vanderbilt.edu/wiki/Main/PowerSampleSize).
Results
Our study included more extrahepatic (EH-CCA:n=141) than intrahepatic CCA (IH-CCA:n=41). As assessed by Student'sttest, there was a significant overrepresentation of female gender in the control cohort (ratio 1.33 in controls versus 0.75 in cancer subjects;P=0.01). Further attesting to the epidemiology of CCA, the median age at study entry was significantly lower in the control group (67.3±11.8 vs 61.3±11.4;P<0.01). Demographic and clinical characteristics of the study groups are shown in Table 1.
Allelic discrimination forrs2910164was successful in all participants (call rate 100%), and re-genotyping in >10% of samples yielded congruent results for each individual. The genotype distribution in the control cohort was consistent with the HWE (P>0.05). As for genotyping results, no significant differences in either allele or genotype frequencies were detected between the CCA and control cohorts. The minor allele frequency was 20.6% in the CCA group compared to 22.3% in the control group. However, there was a non-significant trend for a protective effect of the heterozygous SNP state GC, as indicated by an underrepresentation of heterozygotes in the CCA group [29.1% vs 34.9%; odds ratio (OR)=0.77, 95% confidence interval (95% CI), 0.52-1.13;P=0.18 as per the Chi-square test for 2×2 contingency tables; crude ORs are given]. The allele and genotype distributions ofrs2910164in the study groups is summarized in Table 2. As regards the disparate gender and age structure between the groups, multivariate logistic regression analysis with astepwise forward approach proved age and gender to represent significant and independent factors associated with CCA risk in our sample set. Multivariate analysis incorporatingrs2910164variation was not performed, since, in accord with CCA epidemiology, only age and gender but not genotype were identified as CCA risk factors by univariate tests (for age: OR=1.038; 95% CI, 1.02-1.06;P=0.0001; for gender: OR=0.582; 95% CI,0.41-0.84;P=0.003). It has to be noted, however, that given the non-stringent prospective recruitment of control and CCA individuals in our data set, although consistent with CCA epidemiology in general, these associations could also, at least in part, reflect gender and age disproportions in the study cohort (compared to Table 1). Likewise, exploratory data analyses stratifying according to gender and extra- versus intrahepatic tumor localization did not yield significant associations with CCA risk (Tables 3 and 4). However, the trend towards a protective genotype effect of GC heterozygosity detected in the global analysis was amplified in the EHCCA subgroup, although this marginally fell short of reaching significance (26% vs 35%; OR=0.67; 95% CI, 0.43-1.02;P=0.065). Moreover, we tried to assess the potential influence ofrs2910164on early-onset CCA by arbitrarily limiting our analysis to patients below 60 years at study entry/tumor diagnosis. Similarly, no significant difference was observed between the groups (Table 5).
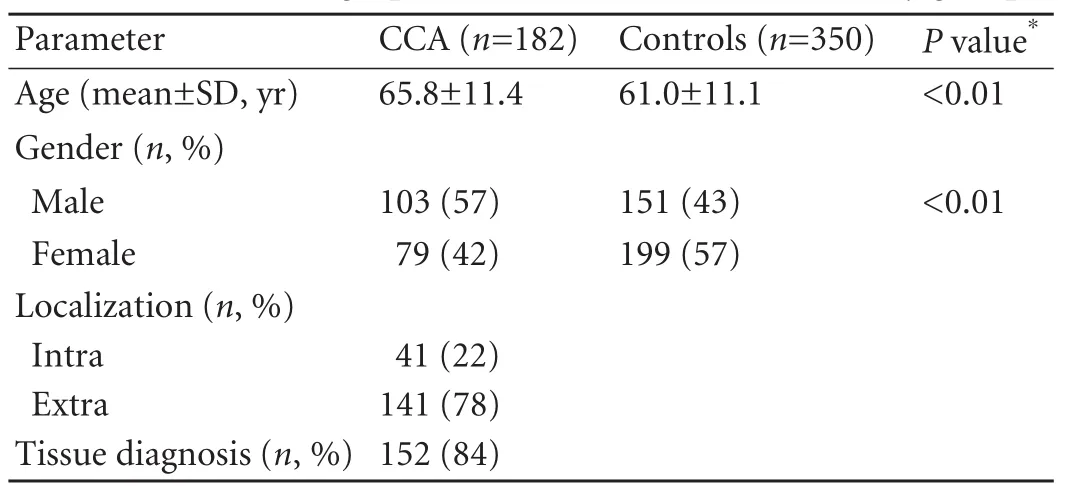
Table 1. Basic demographic and clinical data of the study groups
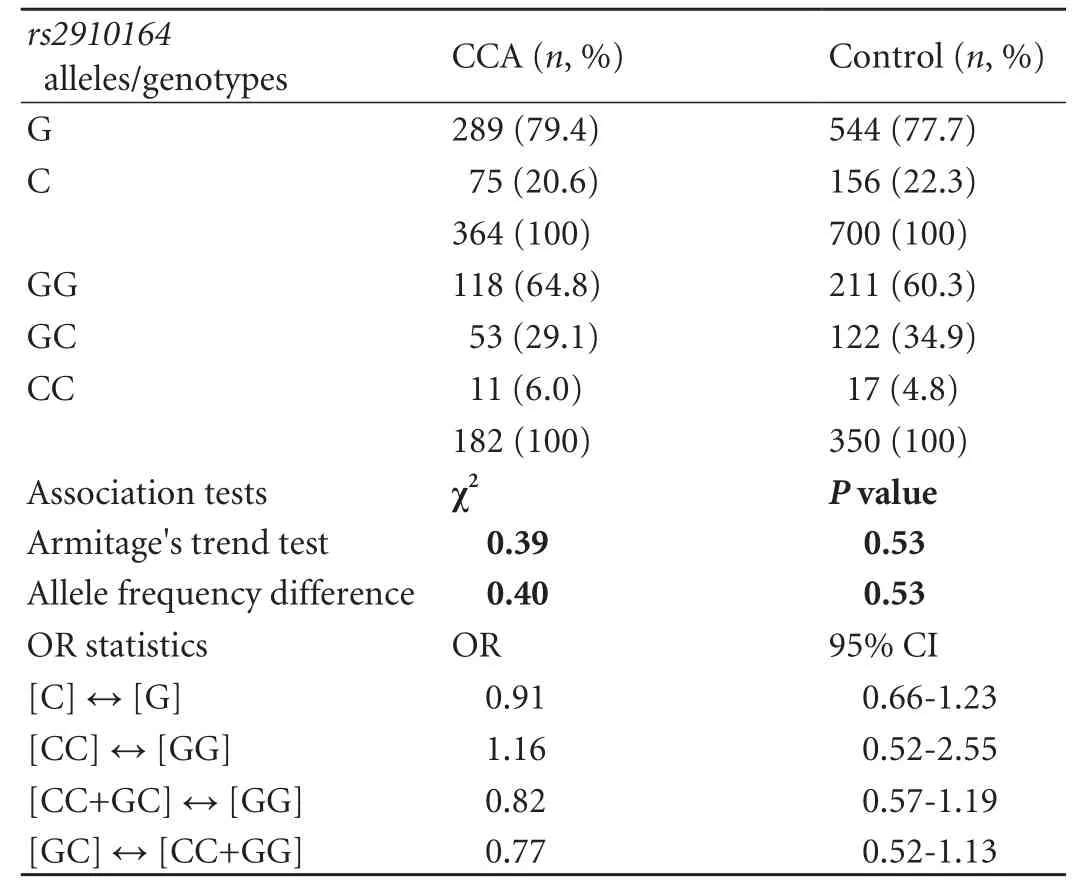
Table 2. Allele and genotype distributions of rs2910164 in the study groups
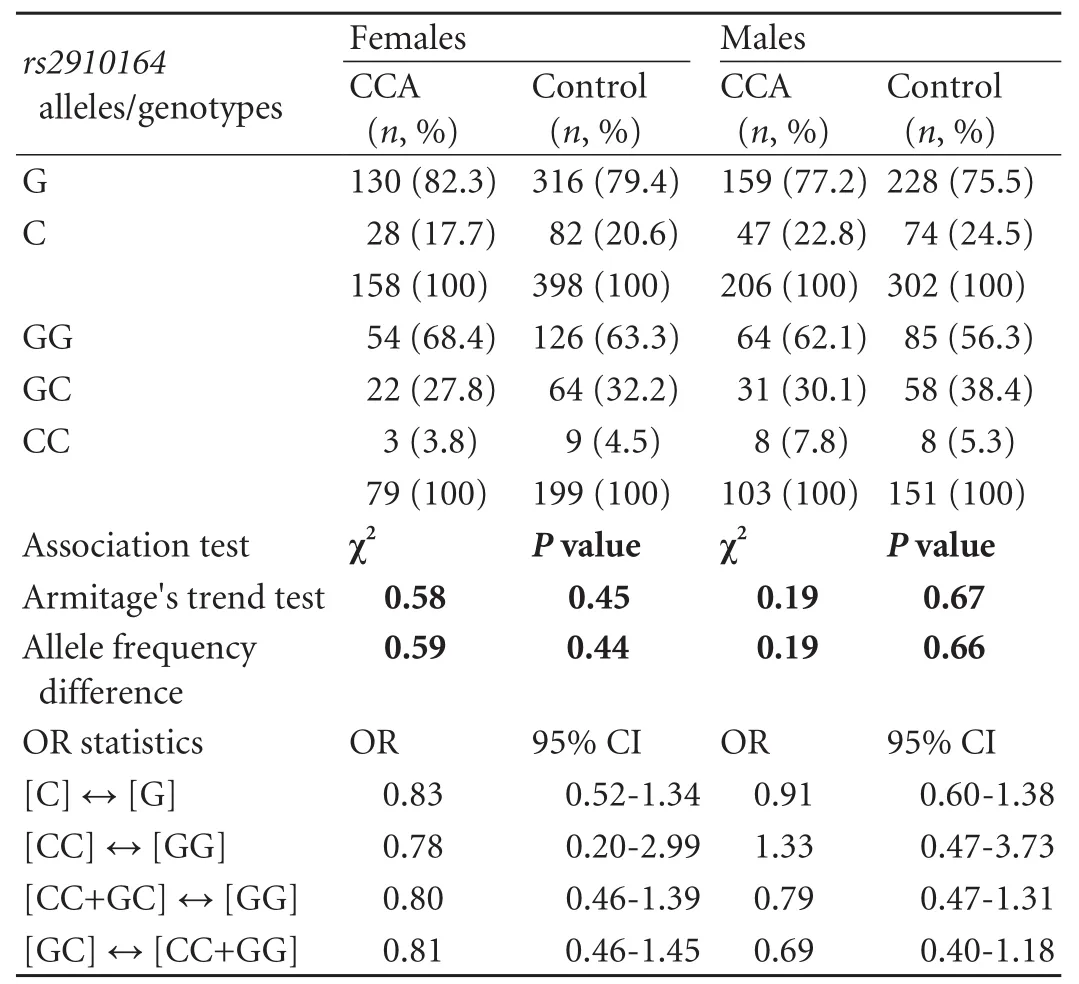
Table 3. Allele and genotype frequencies of rs2910164 in the gender-related subgroups
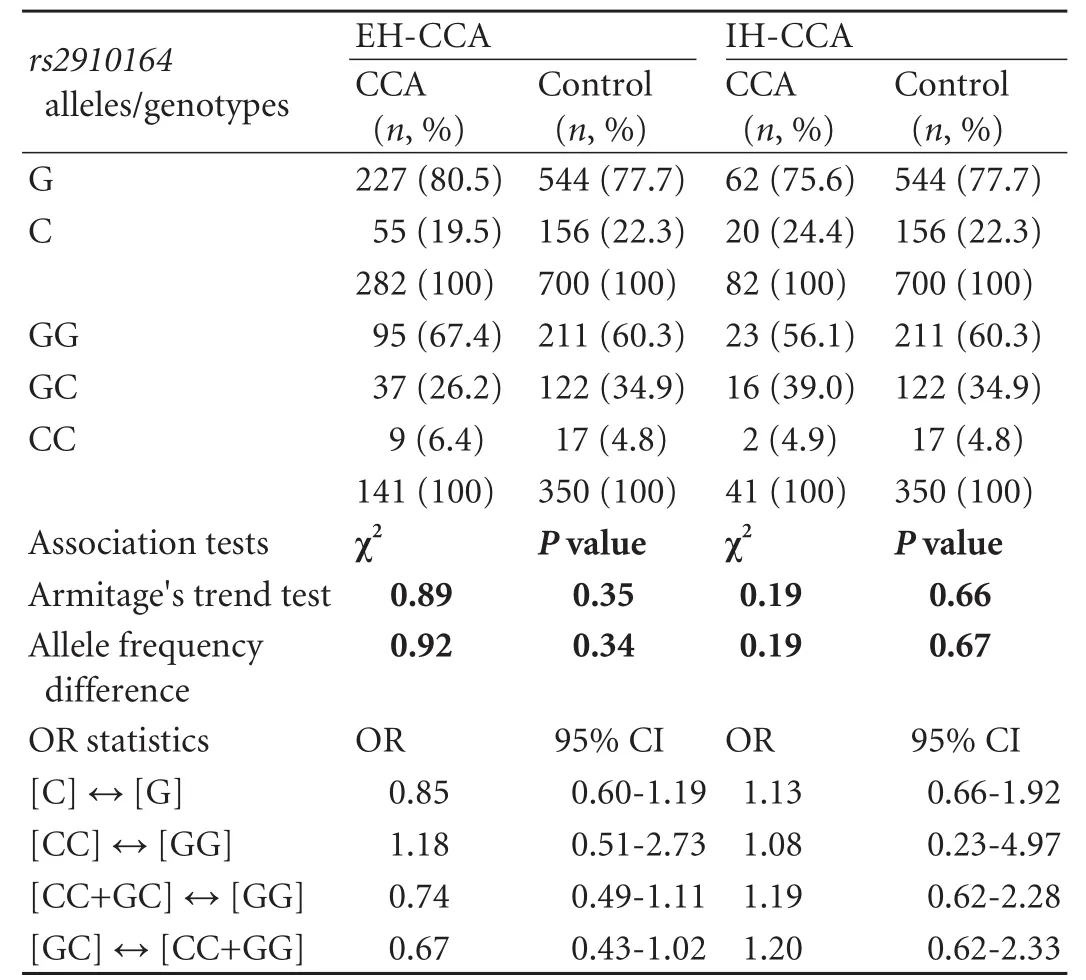
Table 4. Allele and genotype frequencies of rs2910164 in the tumor-stratified subgroups
Thus, in order to assess for a potential type 2 error with respect to our overall negative association, we performed power calculations. The power to identify an effect size of reasonable impact (an OR of 1.7) was calculated at 95.5%, indicating appropriate power to exclude major effects of the pre-miR-146avariant on cholangiocarcinogenesis. Forrs2910164GC heterozygosity, study power was attenuated, with 81.5% power to pinpoint an association signal at an OR of 1.7 (Fig.).
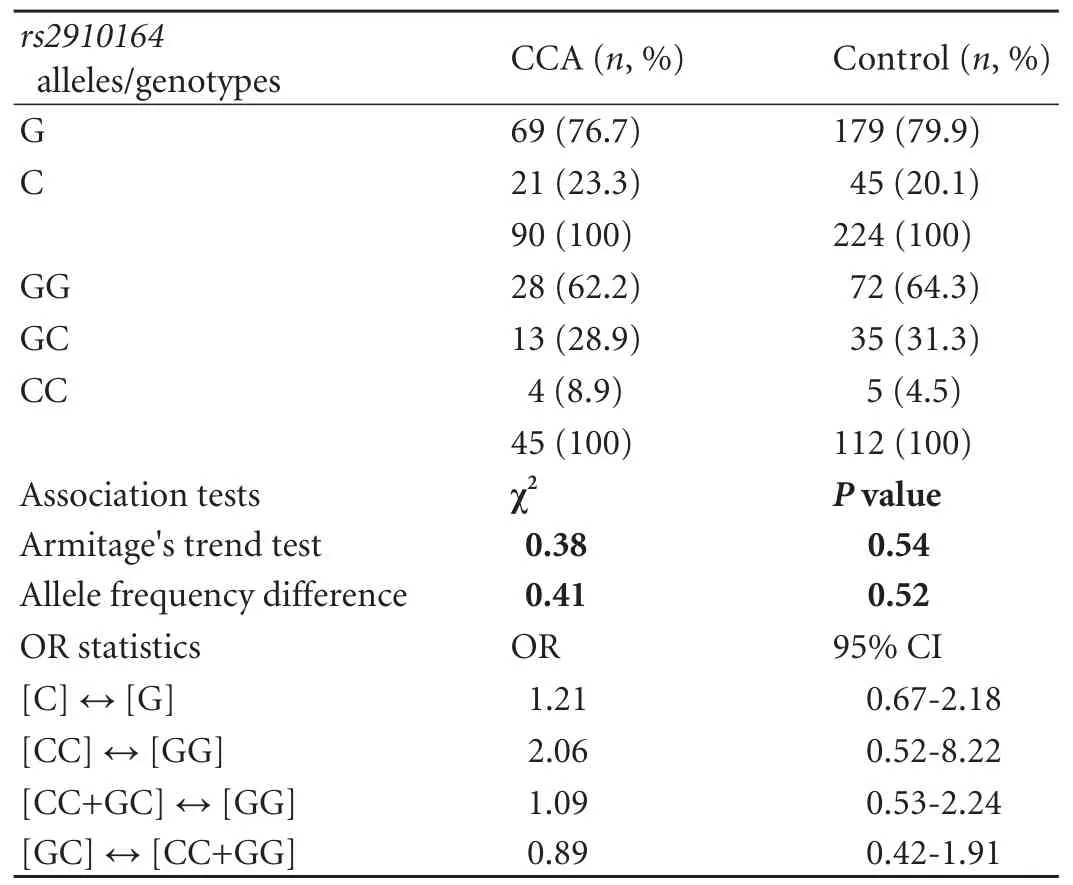
Table 5. Allele and genotype frequencies of rs2910164 in earlyonset CCA

Fig. Statistical power as a function of the effect size (odds ratio) and rs2910164 minor allele frequency (black) or rs2910164 GC heterozygosity rate (grey) (α set at 0.05).
Discussion
The G/C variant in the precursor sequence ofmiR-146alocated in the passenger strand results in mis-pairing of hairpin structure, and has emerged as one of the first miR-SNPs associated with various cancer types.[15]Beyond papillary thyroid cancer and hepatocellular carcinoma,rs2910164has recently been implicated, though with conflicting results, in the genetic risk of gastric carcinoma in Asian populations.[16,17]Moreover, this variant has been associated with susceptibility to esophageal squamous cell carcinoma and head and neck tumors.[18,19]On a molecular level, there are intricate relationships betweenmiR-146aand NF-κB signaling, considered to represent a hallmark pathway in inflammation-driven cancers such as CCA.[20]NF-κBhas been reported to up-regulatemiR-146aexpression after activation of specific toll-like receptors (TLRs) as well as tumor necrosis factor α (TNF-α) and interleukin 1β (IL-1β) stimulation.[6,21]Since the genes of important downstream NF-κB pathway partners including IL-1 receptor-associated kinase 1 (IRAK1) and TNF receptorassociated factor 6 (TRAF6) have been reported to be directmiR-146atargets,miR-146ahas been suggested to provide an important negative regulation of pathogenic NF-κB signaling.[22]
This is the first study to address the potential role ofrs2910164in the genetic predisposition to CCA including 182 individuals from the currently largest Europeanbased CCA cohort. Given the overall negative association, power estimates provided robust statistical power >95% to exclude a reasonably strong allelic effect ofrs2910164with an OR of 1.7. Thus, the study might be considered to be adequately powered to exclude a statistical type 2 error, i.e., a false negative association signal. However, taking into consideration the evidence from association studies of thyroid cancer with a reported implication of pre-miR-146aSNP heterozygosity translating into a distinct repertoire of target genes, we also explored this possibility in the current analysis. Nevertheless, also from the latter perspective, no formally significant indication for increased genetic risk of CCA arose, although it has to be noted that the statistical power to firmly refute this hypothesis was limited compared to the overall study power (82% power for detection of an OR of 1.7). Given the gender differences in CCA epidemiology and the fact that EH- and IH-CCA have been proposed to be different entities on a molecular and histogenetic level, we also performed an explorative data analysis by stratifying according to tumor localization and gender.[9]However, no significant differences in allele and/or genotype distributions emerged. Of interest, pre-miR-146aG/C has been suggested to modulate the age at cancer onset in breast and ovarian cancer patients, while notper sebeing linked to breast cancer risk.[23-25]Taking into account that genetic factors might play a more important role in younger cancer patients, we also studied a subgroup of "early-onset" CCA patients. This was arbitrarily defined as age below 60 years at cancer diagnosis, however, this analysis likewise did not yield significant differences in allele and/or genotype frequencies.
In conclusion, notwithstanding the potential limitation of our study design owing to mismatching of groups in terms of age and gender structure, these data from a large European CCA cohort do not support a prominent contribution of thepre-miR-146aG/C polymorphism to the genetic predisposition to CCA formation. However, given the reduced statistical power with regard to this specific issue, a potential subtle risk modulation by GC heterozygosity may not be excluded and might be evaluated further in larger cohorts.
Contributors:ZV and LF proposed the study. MF, LF and ZV wrote the first draft, and analyzed the data. All authors contributed to the design and interpretation of the study and to further drafts. ZV is the guarantor.
Funding:This study was supported by a grant from the HOMFOR (T201000688 to ZV).
Ethical approval:The local Ethics Committees approved the study, which was in accordance with the revisedDeclaration of Helsinki.
Competing interest:No benefits in any form have been received or will be received from a commercial party related directly or indirectly to the subject of this article.
1 Mott JL. MicroRNAs involved in tumor suppressor and oncogene pathways: implications for hepatobiliary neoplasia. Hepatology 2009;50:630-637.
2 Ryan BM, Robles AI, Harris CC. Genetic variation in microRNA networks: the implications for cancer research. Nat Rev Cancer 2010;10:389-402.
3 Jazdzewski K, Murray EL, Franssila K, Jarzab B, Schoenberg DR, de la Chapelle A. Common SNP in pre-miR-146adecreases mature miR expression and predisposes to papillary thyroid carcinoma. Proc Natl Acad Sci U S A 2008;105:7269-7274.
4 Xu T, Zhu Y, Wei QK, Yuan Y, Zhou F, Ge YY, et al. A functional polymorphism in themiR-146agene is associated with the risk for hepatocellular carcinoma. Carcinogenesis 2008;29:2126-2131.
5 Jazdzewski K, Liyanarachchi S, Swierniak M, Pachucki J, Ringel MD, Jarzab B, et al. Polymorphic mature microRNAs from passenger strand of pre-miR-146acontribute to thyroid cancer. Proc Natl Acad Sci U S A 2009;106:1502-1505.
6 Jazdzewski K, de la Chapelle A. Genomic sequence matters: a SNP in microRNA-146a can turn anti-apoptotic. Cell Cycle 2009;8:1642-1643.
7 Blechacz B, Gores GJ. Cholangiocarcinoma: advances in pathogenesis, diagnosis, and treatment. Hepatology 2008;48: 308-321.
8 Welzel TM, Graubard BI, Zeuzem S, El-Serag HB, Davila JA, McGlynn KA. Metabolic syndrome increases the risk of primary liver cancer in the United States: a study in the SEERMedicare database. Hepatology 2011;54:463-471.
9 Tyson GL, El-Serag HB. Risk factors for cholangiocarcinoma. Hepatology 2011;54:173-184.
10 Höblinger A, Lammert F. Genetics of biliary tract diseases: new insights into gallstone disease and biliary tract cancers. Curr Opin Gastroenterol 2008;24:363-371.
11 Mihalache F, Höblinger A, Grünhage F, Krawczyk M, Gärtner BC, Acalovschi M, et al. Heterozygosity for the alpha1-antitrypsin Z allele may confer genetic risk of cholangiocarcinoma. Aliment Pharmacol Ther 2011;33:389-394.
12 Kawahigashi Y, Mishima T, Mizuguchi Y, Arima Y, Yokomuro S, Kanda T, et al. MicroRNA profiling of human intrahepaticcholangiocarcinoma cell lines reveals biliary epithelial cellspecific microRNAs. J Nihon Med Sch 2009;76:188-197.
13 Karakatsanis A, Papaconstantinou I, Gazouli M, Lyberopoulou A, Polymeneas G, Voros D. Expression of microRNAs, miR-21, miR-31, miR-122, miR-145, miR-146a, miR-200c, miR-221, miR-222, and miR-223 in patients with hepatocellular carcinoma or intrahepatic cholangiocarcinoma and its prognostic significance. Mol Carcinog 2011.
14 Khan SA, Davidson BR, Goldin R, Pereira SP, Rosenber g WM, Taylor-Robinson SD, et al. Guidelines for the diagnosis and treatment of cholangiocarcinoma: consensus document. Gut 2002;51:VI1-9.
15 Fabbri M, Valeri N, Calin GA. MicroRNAs and genomic variations: from Proteus tricks to Prometheus gift. Carcinogenesis 2009;30:912-917.
16 Zeng Y, Sun QM, Liu NN, Dong GH, Chen J, Yang L, et al. Correlation between pre-miR-146aC/G polymorphism and gastric cancer risk in Chinese population. World J Gastroenterol 2010;16:3578-3583.
17 Okubo M, Tahara T, Shibata T, Yamashita H, Nakamura M, Yoshioka D, et al. Association between common genetic variants in pre-microRNAs and gastric cancer risk in Japanese population. Helicobacter 2010;15:524-531.
18 Liu Z, Li G, Wei S, Niu J, El-Naggar AK, Sturgis EM, et al. Genetic variants in selected pre-microRNA genes and the risk of squamous cell carcinoma of the head and neck. Cancer 2010;116:4753-4760.
19 Guo H, Wang K, Xiong G, Hu H, Wang D, Xu X, et al. A functional varient in microRNA-146a is associated with risk of esophageal squamous cell carcinoma in Chinese Han. Fam Cancer 2010;9:599-603.
20 Karin M, Greten FR. NF-kappaB: linking inflammation and immunity to cancer development and progression. Nat Rev Immunol 2005;5:749-759.
21 Li L, Chen XP, Li YJ. MicroRNA-146a and human disease. Scand J Immunol 2010;71:227-231.
22 Taganov KD, Boldin MP, Chang KJ, Baltimore D. NF-kappaB-dependent induction of microRNA miR-146, an inhibitor targeted to signaling proteins of innate immune responses. Proc Natl Acad Sci U S A 2006;103:12481-12486.
23 Shen J, Ambrosone CB, DiCioccio RA, Odunsi K, Lele SB, Zhao H. A functional polymorphism in themiR-146agene and age of familial breast/ovarian cancer diagnosis. Carcinogenesis 2008;29:1963-1966.
24 Pastrello C, Polesel J, Della Puppa L, Viel A, Maestro R. Association between hsa-miR-146agenotype and tumor ageof-onset in BRCA1/BRCA2-negative familial breast and ovarian cancer patients. Carcinogenesis 2010;31:2124-2126.
25 Gao LB, Bai P, Pan XM, Jia J, Li LJ, Liang WB, et al. The association between two polymorphisms in pre-miRNAs and breast cancer risk: a meta-analysis. Breast Cancer Res Treat 2011;125:571-574.
December 30, 2011
Accepted after revision March 28, 2012
Author Affiliations: Department of Medicine II, Saarland University Hospital, 66421 Homburg, Germany (Mihalache F, Lammert F and Zimmer V); Department of Medicine III, University Iuliu Hatieganu, Cluj-Napoca, Romania (Mihalache F and Acalovschi M); Department of Internal Medicine I, University Hospital Bonn, Bonn, Germany (Höblinger A and Sauerbruch T)
Vincent Zimmer, MD, Department of Medicine II, Saarland University Hospital, Homburg 66421, Germany (Tel: +49-6841-1623221; Fax: +49-6841-1623264; Email: vincent.zimmer@uks.eu)
© 2012, Hepatobiliary Pancreat Dis Int. All rights reserved.
10.1016/S1499-3872(12)60200-8
杂志排行

Hepatobiliary & Pancreatic Diseases International的其它文章
- Gastric- and intestinal-type marker expression in invasive ductal adenocarcinoma of the pancreas
- Expression of SOCS3 throughout liver regeneration is not regulated by DNA methylation
- Early changes of hepatic hemodynamics measured by functional CT perfusion in a rabbit model of liver tumor
- Muscarinic acetylcholine receptor M3 in proliferation and perineural invasion of cholangiocarcinoma cells
- Hepatocyte differentiation of mesenchymal stem cells
- Early control of short hepatic portal veins in isolated or combined hepatic caudate lobectomy