猪瘟病毒和不同型猪繁殖与呼吸综合征病毒的多重RT-PCR检测方法的建立
2012-06-29莫胜兰施开创邹联斌陆文俊
莫胜兰,施开创,胡 杰,邹联斌,陆文俊,李 军
(广西动物疫病预防控制中心,广西 南宁 530001)
猪繁殖与呼吸综合征病毒(Porcine reproduc-tive and respiratory syndrome virus,PRRSV)可以引起以母猪繁殖障碍和各种年龄猪(特别是仔猪)严重的呼吸道疾病为主要特征的猪繁殖与呼吸综合征(PRRS)。PRRSV可以分为两种基因型,即以VR-2332株为代表的美洲型毒株和以Lelystad virus(LV)为代表的欧洲型毒株,两者之间的全基因组核苷酸序列同源性仅为60%左右。我国于1996年首次分离到PRRSV美洲型经典毒株[1],2006年分离到以Nsp2蛋白发生第481位氨基酸和第533~561位氨基酸30个氨基酸缺失为标志的高致病性美洲型变异毒株(HP-PRRSV)[2],2010年分离到致病性PRRSV欧洲型毒株[3]。猪瘟病毒(Classical swine fever virus,CSFV)可以引起各种年龄猪以高热稽留、全身广泛性出血以及母猪繁殖障碍为主要特征的猪瘟(CSF)。CSFV属于单股正链RNA病毒,基因组大约12.3kb,含有一个大的编码3898氨基酸的阅读框(ORF)。CSF和PRRS均为我国一类动物疫病,在我国各地时有发生,并且存在病原隐性感染和持续性感染的现象[4-5]。同时 CSFV 和PRRSV混合感染和继发感染严重[6],给养猪业造成巨大的经济损失,严重危害我国养猪业的健康发展。目前,已建立了检测CSFV[7-8]、PRRSV[9-10]的单一RT-PCR方法,也建立了检测CSFV 和PRRSV的多重 RT-PCR方法[11-12],但迄今未见报道能同时检测并区分CSFV以及PRRSV美洲型经典毒株、变异毒株和欧洲型毒株的RT-PCR方法。本研究针对CSFV、PRRSV的基因序列设计3对特异性引物,建立了能同时检测并区分CSFV以及PRRSV美洲型经典毒株、变异毒株和欧洲型毒株的多重RT-PCR方法,为CSFV和PRRSV的快速检测以及流行病学研究提供了有效的技术平台。
1 材料与方法
1.1 材料
1.1.1 病毒及重组质粒 PRRSV美洲型经典毒株(VR-2332株)、美洲型变异毒株(疫苗毒JXA1-R株)、猪瘟病毒(CSFV,疫苗毒C株)、猪伪狂犬病病毒(PRV,疫苗毒 Bartha-K61 株)、猪细小病毒(PPV,疫苗毒N株)、猪口蹄疫病毒(FMDV,疫苗毒 OS/99株)、猪圆环病毒2型(PCV-2,疫苗毒 LG株)由广西动物疫病预防控制中心实验室保存。含有PRRSV美洲型毒株ORF5基因的重组质粒p-AM-ORF5、分别含有欧洲型毒株Nsp2基因片段、ORF567基因片段的重组质粒p-EU-ORF1a、p-EUORF567由中国农业科学院北京畜牧兽医研究所兽医研究室惠赠。
1.1.2 主要试剂 MiniBEST Viral RNA/DNA Extraction Kit Ver.4.0试剂盒、RNA PCR Kit(AMV)Ver.3.0试剂盒、Agarose Gel DNA Purification Kit Ver.2.0试剂盒、pMD18-T 载体、质粒DNA小量提取试剂盒为宝生物工程(大连)有限公司产品;DH5α感受态细胞为北京天根生化科技有限公司产品。
1.2 方法
1.2.1 引物设计与合成 参考GenBank上登录的CSFV石门株(AF092448)、PRRSV美洲型经典毒株VR-2332株(AY150564)、高致病性变异毒株JXA1株(EF112445)以及欧洲型毒株LV株(M96262)的基因组序列,设计3对特异性引物(表1)。其中,CSF-NS2引物对扩增CSFV NS2基因片段的大小为508bp;AM-Nsp2引物对扩增PRRSV美洲型毒株Nsp2基因片段的大小在经典毒株为338bp、在变异毒株为248bp;EU-ORF5引物对扩增PRRSV欧洲型毒株ORF5基因片段的大小为614bp。引物由宝生物工程(大连)有限公司合成。
1.2.2 病毒重组质粒标准品制备 分别取CSFV C株和 PRRSV VR-2332株、JXA1-R 株细胞培养液,用RNA提取试剂盒提取病毒总RNA后,应用PCR试剂盒进行反转录获得cDNA。以病毒cDNA或重组质粒p-EU-ORF567为模板,分别应用3对特异性引物,建立50μL PCR反应体系:5×buffer 10μL,灭菌蒸馏水33.75μL,Ex Taq HS聚合酶(5 U/μL)0.25μL,上、下游引物(25pmol/μL)各0.5 μL,cDNA 5μL。反应程序为:94℃2min;94℃30s,56℃~58℃30s,72℃45s,35个循环;最后72℃10min。回收PCR产物、连接pMD18-T载体、转化到DH5α感受态细胞、培养后提取质粒进行PCR、酶切及测序鉴定。本研究共构建4种重组质粒,其中由引物CSF-NS2扩增产物构建的重组质粒命名为p-CSF-NS2,引物 AM-Nsp2扩增产物构建的重组质粒分别命名为 p-Nsp2-VR2332、p-Nsp2-JXA1,引物EU-ORF5扩增产物构建的重组质粒命名为p-EU-ORF5。

表1 多重RT-PCR特异性引物Table1 The specific primers used for the multiplex RT-PCR
1.2.3 多重PCR退火温度的确定 建立50μL PCR反应体系:5× RT buffer 10μL,Ex Taq HS聚合酶(5U/μL)0.25μL,3对特异性引物的上、下游引物(25pmol/μL)各0.5μL,等比例混合的4种重组质粒5μL,灭菌双蒸水补至50μL。在55.1℃~60℃间设定6个退火温度进行梯度PCR,确定最佳退火温度。
1.2.4 多重PCR引物浓度的确定 50μL PCR反应体系中,3对引物终浓度分别为0.5、0.75、1.0、1.25、1.5pmol/μL,进行不同浓度的排列组合,进行PCR,确定最佳引物浓度。
1.2.5 多重PCR聚合酶浓度的确定 50μL PCR反应体系中,Ex Taq HS聚合酶(5U/μL)终浓度分别为0.015、0.02、0.025、0.03、0.035U/μL,进行PCR,确定最佳聚合酶浓度。
1.2.6 多重PCR循环数的确定 建立50μL PCR反应体系,分别设置30、35、40、45个循环,进行PCR,确定最佳循环数。
1.2.7 多重PCR的特异性试验 以重组质粒p-CSF-NS2、p-Nsp2-VR2332、p-Nsp2-JXA1、p-EUORF5,以及FMDV的cDNA和PCV-2、PRV、PPV的DNA作为模板,在优化后的反应条件下进行多重PCR,分析其特异性。
1.2.8 多重PCR的敏感性试验 将4种重组质粒10倍系列稀释成1.67×1010拷贝/μL~1.67×101拷贝/μL(50μL反应体系终浓度为1.67×109拷贝/μL~1.67×100拷贝/μL)共10个浓度梯度。将同一浓度梯度的各种质粒等体积混合后作为模板,在优化后的反应条件下进行多重PCR,分析其敏感性。
1.2.9 多重PCR的重复性试验 以等体积混合的4种重组质粒为模板,在优化后的反应条件下进行多重PCR,重复5次反应,分析其重复性。
1.2.10 多重PCR的临床应用 临床病料来自2010年-2011年广西各地发病死亡的疑似病猪,共106份。取脾、肺、淋巴结各约5g,混合研磨后按1∶4(W/V)加入 PBS(pH7.2),反复冻融3次,10000r/min离心10min,取上清提取总RNA。应用所建立的多重 RT-PCR方法,进行CSFV和PRRSV检测。
2 结果
2.1 重组质粒的鉴定
分别以CSFV C株和PRRSV VR-2332株、JXA1-R株的cDNA以及欧洲型毒株重组质粒p-EU-ORF567为模板进行PCR,扩增相应的目的片段,经回收、连接、转化、鉴定,成功构建了重组质粒p-CSF-NS2、p-Nsp2-VR2332、p-Nsp2-JXA1 和 p-EU-ORF5(图1)。
2.2 最佳退火温度的确定
设定6个退火温度进行梯度PCR,各个退火温度均能扩增出条带(图2)。在57.3℃条带最清晰,且不存在非特异性条带,确定为最佳退火温度。
2.3 最佳引物浓度的确定
建立50μL反应体系,固定其它条件不变,以3对引物CSF-NS2、AM-Nsp2及EU-ORF5不同浓度的排列组合进行多重PCR。最终确定各对引物的最佳浓度(反应终浓度):CSF-NS2为0.5pmol/μL、AMNsp2为1.25pmol/μL、EU-ORF5为0.5pmol/μL(图3)。
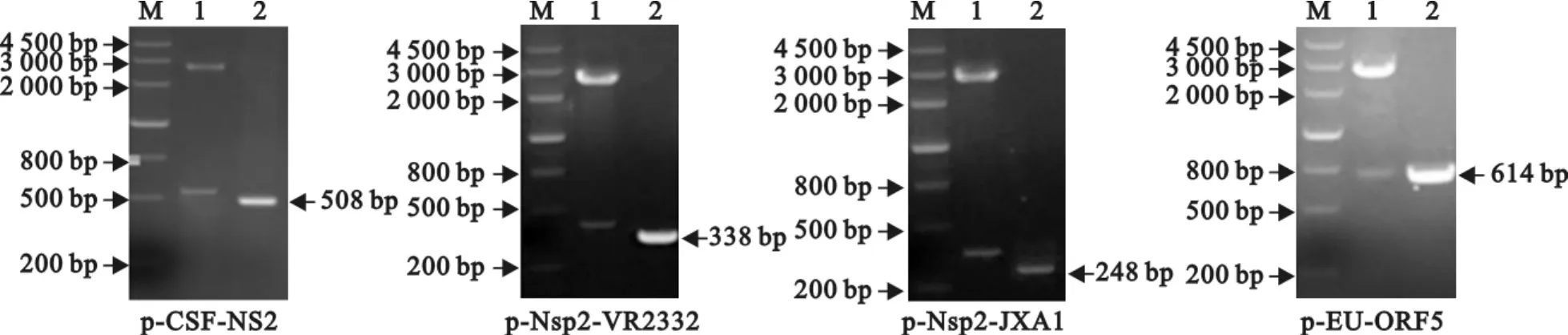
图1 重组质粒鉴定Fig.1 Identification of the recombinant plasmids
2.4 最佳聚合酶浓度的确定
建立50μL反应体系,固定其它条件不变,Taq HS聚合酶分别以0.0125、0.02、0.025、0.03、0.035 U/μL浓度进行多重PCR,最终确定最佳聚合酶浓度为0.03U/μL(图4)。
2.5 最佳循环数的确定
建立50μL反应体系,固定其它条件不变,分别设置30、35、40、45个循环进行PCR,最终确定最佳循环数为40个(图5)。
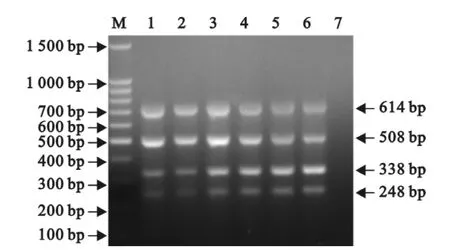
图2 退火温度的选择Fig.2 Selection of the annealing temperature for the multiplex RT-PCR
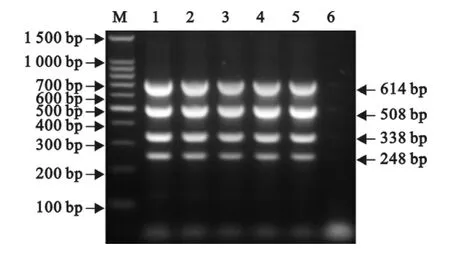
图3 引物浓度的选择Fig.3 Selection of the primer concentration for the multiplex RT-PCR
2.6 最佳反应条件的确定
经优化,多重RT-PCR包括反转录和PCR扩增两步程序。10μL反转录体系为:无RNA酶灭菌双蒸水 0.75μL,10× RT buffer[100mmol/L Tris-HCl(pH8.3),500mmol/L KCl]1.0μL,MgCl2(25mmol/L)2.0μL,dNTP(各10mmol/L)1.0μL,随机引物9聚体(50pmol/μL)0.5μL,RNA酶抑制剂(40U/μL)0.25μL,AMV反转录酶(5U/μL)0.5μL,模板总RNA 4.0μL;反转录程序为:30℃10min,42℃30min,99℃5min。50μL PCR反应体系为:5×RT buffer 10μL,Ex Taq HS聚合酶(5U/μL)0.3 μL,引物CSF-NS2(25 pmol/μL)各1μL、引物 AM-Nsp2(25pmol/μL)各2.5μL、引物 EU-ORF5(25pmol/μL)各1μL,cDNA 5μL,灭菌双蒸水补至50μL;反应程序为:94℃2min;94℃30s,57.3℃30s,72℃45s,40个循环;72℃10min。
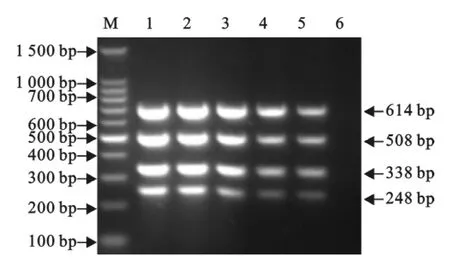
图4 聚合酶浓度的选择Fig.4 Selection of the Taq polymerase concentration for the multiplex RT-PCR
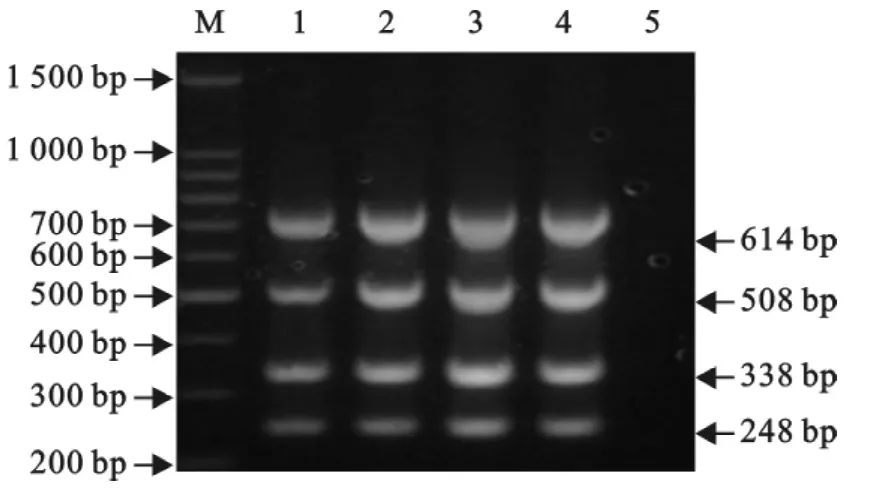
图5 循环数的选择Fig.5 Selection of the reaction cycles for the multiplex RT-PCR
2.7 特异性分析
以4种重组质粒的不同排列组合,以及FMDV的cDNA和PCV-2、PRV、PPV的DNA作为模板,进行多重PCR。结果只有4种重组质粒的不同排列组合反应呈阳性,而 FMDV、PCV-2、PRV、PPV均为阴性,没有交叉反应(图6)。
2.8 敏感性分析
分别以10倍系列稀释的4种重组质粒为模板进行单重 PCR。结果,引物 CSF-NS2以 p-CSFNS2为模板的检出下限为1.67×103拷贝/μL,引物AM-Nsp2以质粒p-Nsp2-VR2332 或 p-Nsp2-JXA1为模板的检出下限均为1.67×102拷贝/μL,引物EU-ORF5以质粒p-EU-ORF5为模板的检出下限为1.67×103拷贝/μL(图7)。而将10倍系列稀释的4种重组质粒等体积混合后作为模板进行多重PCR,结果3对引物的检出下限均为1.67×103拷贝/μL(图8)。
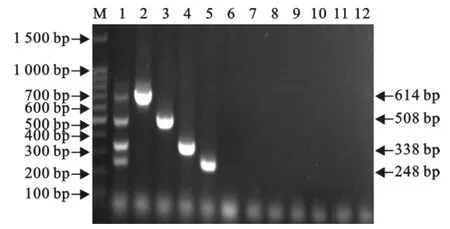
图6 多重PCR的特异性试验Fig.6 The specificity test of the multiplex RT-PCR

图7 PCR的敏感性试验Fig.7 The sensitivity test of the single RT-PCR
2.9 重复性分析
以含有1.67×108拷贝/μL重组质粒混合物为模板,在优化后的反应条件下进行多重PCR。结果显示,5次重复试验均能扩增出均匀一致的目的片段(图9),具有很好的重复性。
2.10 临床病料检测结果
应用所建立的多重RT-PCR方法对106份临床病料进行检测。结果CSFV和PRRSV变异株混合感染阳性病科4份,占3.77%(4/106);CSFV阳性7份,占6.60%(7/106);PRRSV变异株阳性17份,占16.04%(17/106)。随机选取上述阳性样品各4份,回收PCR产物、克隆、测序,证实为CSFV或PRRSV。部分临床病料检测结果见图10。
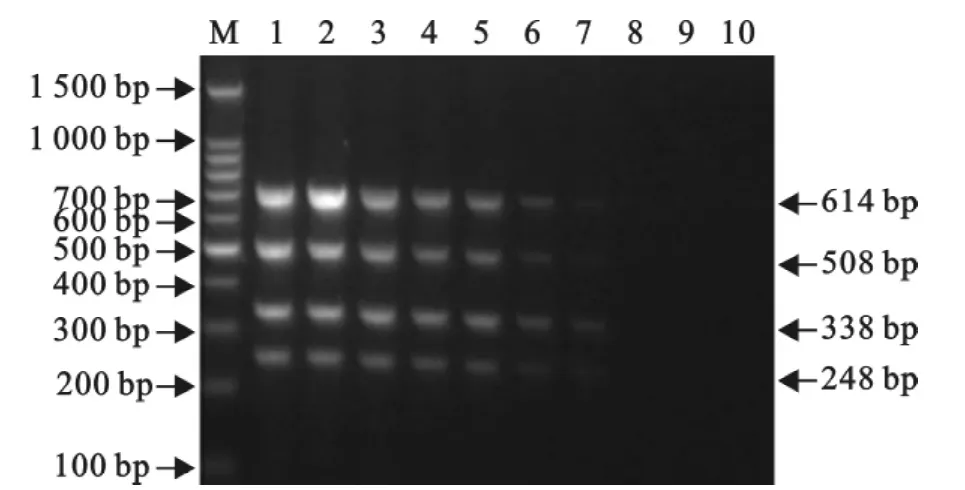
图8 多重PCR的敏感性试验Fig.8 The sensitivity test of the multiplex RT-PCR
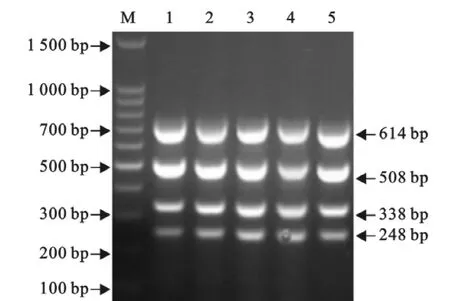
图9 多重PCR的重复性试验Fig.9 The repeatability test of the multiplex RT-PCR

图10 部分临床样品的检测结果Fig.10 The detection results of the clinical samples by the multiplex RT-PCR
3 讨论
当前,PRRSV美洲型经典毒株、变异毒株(HPPRRSV)以及欧洲型毒株在我国均有流行,HPPRRSV仍然是我国的优势流行毒株,而新近证实在我国存在的欧洲型毒株间也具有明显的基因组差异[13],因此PRRSV流行毒株在临床上日益复杂。猪瘟是我国猪群的老疫病,经过多年采取强制免疫、扑杀及综合防控措施,该病得到有效控制。当前,全世界的CSFV流行毒株可以分为3个基因群,在我国流行的毒株为基因1群和2群,其中基因1群与我国1945年分离的石门强毒有很近的遗传关系,在我国猪瘟流行中占次要地位;基因2群与欧洲流行的基因2群病毒有较近遗传关系,在我国猪瘟流行中占主导地位[14]。HP-PRRSV和CSFV已成为近年来严重危害我国猪群健康的两种重要病原,而且PRRSV和CSFV继发感染、混合感染屡见不鲜[6],由此给养猪业造成的损失日益严重。因此,建立能同时检测并区分CSFV以及PRRSV美洲型经典毒株、HP-PRRSV、欧洲型毒株的病原学诊断方法,对快速鉴别诊断CSF和PRRS,以便果断、有效处置疫情意义重大。
本研究根据CSFV基因组序列在NS2基因比较保守[15],以及PRRSV 基因组序列在 Nsp2及ORF5基因变异较大的特点[13,16],利用位于 CSFV NS2基因区域的一对引物扩增CSFV,利用位于PRRSV Nsp2基因高变区的一对引物特异扩增并区分美洲型经典毒株与变异毒株,利用位于ORF5基因高变区的一对引物特异扩增欧洲型毒株,实现了利用3对特异引物在同一个扩增反应中,鉴别4种不同毒株即CSFV以及PRRSV美洲型经典毒株、HP-PRRSV、欧洲型毒株。通过对影响多重RT-PCR反应的主要因素的优化,确定了引物退火温度、引物浓度、聚合酶浓度、反应循环数,获得了多重PCR的最佳反应条件,成功建立了特异性强、敏感性高、重复性好的PRRSV多重RT-PCR检测方法。并且,应用该法检测了106份临床病料,进一步证实了其可靠性。
本研究应用所建立的多重RT-PCR方法检测2010年-2011年采自广西各地的疑似病料,发现CSFV、PRRSV存在单独感染和混合感染现象。在检测的106份病料中,CSFV的阳性率为10.38%(11/106),表明由CSFV给猪群带来的危害依然严重;PRRSV 的阳性率为19.81%(21/106),均为HP-PRRSV,说明PRRSV变异毒株已成为广西流行的优势毒株;虽然未检测到欧洲型毒株阳性病料,但最近国内已分离到致病性欧洲型毒株[3],广西猪群依然面临来自欧洲型毒株的潜在威胁。值得注意的是,CSFV 和PRRSV混合感染占3.77%(4/106),其危害性不容忽视。由于PRRSV感染可以抑制动物机体对猪瘟疫苗的免疫反应,延缓和降低细胞免疫和体液免疫强度[17],导致PRRSV感染猪群即使进行了猪瘟弱毒疫苗的免疫预防接种,但免疫效果不佳,猪群得不到有效的免疫保护,仍然存在猪瘟野毒感染和发病的危险,这可能是近年来一些猪群免疫接种猪瘟疫苗后依然散发猪瘟的原因之一。当前,抓好CSFV和PRRSV的病原学监测和分子流行病学调查,是有效应对日益复杂的CSF和PRRS疫情的重要基础。
本研究针对CSFV和PRRSV基因组序列特点设计3对特异性引物,经过反应条件的优化,成功建立了能够同时检测并区分CSFV以及PRRSV美洲型经典毒株、变异毒株和欧洲型毒株的多重RTPCR方法,为CSFV和PRRSV的快速鉴别诊断和流行病学调查提供了有效的技术平台。
[1]郭宝清,陈章水,刘文兴,等.从疑似PRRS流产胎儿分离PRRSV的研究[J].中国畜禽传染病,1996(2):1-4.
[2]Tian K,Yu X,Zhao T,et al.Emergence of fatal PRRSV variants:unparalleled outbreaks of atypical PRRS in China and molecular dissection of the unique hallmark[J].PloS One,2007,2(6):e526.
[3]Chen N,Cao Z,Yu X,et al.Emergence of novel European genotype porcine reproductive and respiratory syndrome virus in mainland China[J].J Gen Virol,2011,92(Pt4):880-892.
[4]Mills R W,Doster A R,Galeota J A,et al.Duration of infection and proportion of pigs persistently infected with porcine reproductive and respiratory syndrome virus[J].J Clin Microbiol,2003,41(1):58-62.
[5]姚文生,范学政,王 琴,等.我国猪瘟流行现状与防控措施建议[J].中国兽药杂志,2011,45(9):44-47.
[6]王连想,马静云,谢青梅,等.猪繁殖与呼吸综合征病毒变异株与猪瘟病毒等混合感染的调查与分析[J].中国兽医杂志,2009,45(10):3-5.
[7]Díaz de Arce H,Nuñez J I,Ganges L,et al.An RT-PCR assay for the specific detection of classical swine fever virus in clinical samples[J].Vet Res,1998,29(5):431-440.
[8]罗廷荣,莫 扬,吴文德,等.RT-PCR技术检测猪瘟的应用研究[J].中国预防兽医学报,2003,25(3):219-222.
[9]Oleksiewicz M B,Botner A,Madsen K G,et al.Sensitive detection and typing of porcine reproductive and respiratory syndrome virus by RT-PCR amplification of whole viral genes[J].Vet Microbiol,1998,64(1):7-22.
[10]肖跃强,管 宇,唐 娜,等.PRRSV经典与高致病性毒株RT-PCR鉴别检测方法的建立[J].中国兽医学报,2010,30(7):873-877.
[11]孔繁德,王 荣,陈 琼,等.多重RT-PCR一步法技术同时检测猪瘟病毒和蓝耳病病毒方法的建立以及初步应用[J].生物技术通报,2009(1):112-116.
[12]Liu S,Zhao Y,Hu Q,et al.A multiplex RT-PCR for rapid and simultaneous detection of porcine teschovirus,classical swine fever virus,and porcine reproductive and respiratory syndrome virus in clinical specimens[J].J Virol Methods,2011,172(1-2):88-92.
[13]Darwich L,Gimeno M,Sibila M,et al.Genetic and immunobiological diversities of porcine reproductive and respiratory syndrome genotype I strains[J].Vet Microbiol,2011,150(1-2):49-62.
[14]王 琴.我国猪瘟的分子流行病学监测及防控[J].猪业科学,2010(1):82-84.
[15]Leifer I,Hoffmann B,Höper D,et al.Molecular epidemiology of current classical swine fever virus isolates of wild boar in Germany[J].J Gen Virol,2010,91(Pt 11):2687-2697.
[16]Murtaugh M P,Stadejek T,Abrahante J E,et al.The everexpanding diversity of porcine reproductive and respiratory syndrome virus[J].Virus Res,2010,154(1-2):18-30.
[17]Suradhat S,Kesdangsakonwut S,Sada W,et al.Negative impact of porcine reproductive and respiratory syndrome virus infection on the efficacy of classical swine fever vaccine[J].Vaccine,2006,24(14):2634-2642.
