结直肠癌缺失基因功能区诱导结肠癌细胞凋亡
2012-06-28张旭浩焦海展吴亚冉张凯峰门丽慧巩春玲丛中一
张旭浩 焦海展 吴亚冉 张凯峰 门丽慧 赵 青 韩 博 巩春玲 丛中一
(吉林大学药学院再生医学系,吉林 长春 130021)
调查结果表明老年人(≥65岁)占结直肠癌病人的54.00%,老年人直肠癌和结肠癌发病率分别为49.81/10万和63.35/10万,为结直肠癌的高发人群〔1〕,所以结直肠癌的预防和治疗对于人类的健康特别是对于老年人至关重要。研究表明,结直肠癌缺失(DCC)基因的缺失可以导致肿瘤的发生,而DCC基因的表达在诱导肿瘤细胞凋亡方面具有重要作用;同时,有报道17p和18q基因在远地转移肿瘤中缺失的几率很高且可以看到多对等位基因片段的缺失〔2〕。本实验主要研究DCC基因对结肠癌细胞的凋亡诱导作用。
1 材料与方法
1.1 材料 氨苄青霉素、卡那霉素购自北京华美生物工程公司;细菌培养用胰蛋白胨及酵母提取物购自上海生工生物工程技术服务有限公司;细胞培养用胎牛血清(FBS)、DMEM培养基和胰蛋白酶购自GIBCO公司;Lipofectamine 2000购自Invitrogen公司;结肠癌细胞SW1116由本室保存;pIRES2-EGFP质粒以及含有DCC基因胞内功能域(3 727~3 792 bp)的重组表达载体pIRES2-EGFP-DCC为本实验室保存;TUNEL试剂盒购自碧云天生物技术研究所。
1.2 细胞培养 将SW1116细胞在37℃、5%CO2条件下培养于DMEM培养基中(含有10%FBS、100 U/ml青/链霉素),0.25%胰酶消化、传代。
1.3 pIRES2-EGFP-DCC脂质体法转染SW1116细胞 将SW1116细胞按5×103/孔接种于96孔板,分为3组(实验组、质粒对照组、细胞对照组),每组6个平行孔,待细胞贴壁并生长至占板底面积40% ~50%,将4μl Lipofectamine 2000与96μl无双抗无血清培养基混合,室温孵育5 min。然后将pIRES2-EGFP-DCC与pIRES2-EGFP各2μl与48μl无双抗无血清培养基混合,分别加入上述 Lipofectamine 2000稀释液50μl,混合,37℃孵育10 min。实验组96孔板每孔加入 Lipofectamine 2000和pIRES2-EGFP-DCC混合液10μl,24孔板每孔加入100μl;质粒对照组96孔板每孔加入Lipofectamine 2000和pIRES2-EGFP混合液10μl,24孔板每孔加入100μl。5 h后换无双抗有血清培养液,细胞对照组不做处理。
1.4 四甲基耦氮唑盐(MTT)比色法检测转染后细胞增殖情况转染36 h后,于每孔加入10μl MTT(5 mg/ml)。于37℃孵育4 h后弃掉培养液,加入150μl DMSO,摇床震荡15 min,酶标仪检测490 nm每孔吸光度OD值。记录数据并计算细胞生长抑制率,抑制率=(1-实验组OD值/细胞对照组OD值)×100%。
1.5 吖啶橙/溴乙啶(AO/EB)染色法检测转染细胞凋亡情况转染36 h后,于每孔加入AO、EB的等体积混合液2μl,室温避光染色10 min,激发波长510 nm,荧光显微镜下照相观察。镜下随机取5个不重复的视野,计数200个细胞中凋亡及坏死细胞,分别计算细胞凋亡率及坏死率。
1.6 原位末端标记法(TUNEL)检测转染细胞凋亡情况 转染36 h后,方法按稍加修改的TUNEL检测试剂盒说明书进行,使用TUNEL原位检测试剂盒对凋亡细胞进行原位染色。
1.7 统计学处理 应用SPSS11.5统计软件进行数据处理,实验数据以x±s表示,参数统计采用异方差t检验。
2 结果
2.1 MTT比色法检测转染细胞增殖情况 实验组、细胞对照组、质粒对照组的细胞增殖分别为0.599±0.037,0.735±0.031,0.675±0.020,可见转染 pIRES2-EGFP-DCC 36 h后,MTT结果显示,转染DCC基因功能区对SW1116细胞的增殖具有显著的抑制作用,抑制率为32.1%,三组之间的差异有统计学意义(P<0.05)。
2.2 AO/EB染色法检测转染细胞凋亡情况 转染pIRES2-EGFP-DCC 36 h后,AO/EB荧光染色结果如图1所示,实验组中可见大量激发黄色荧光的凋亡晚期细胞,而质粒对照组和细胞对照组中基本没有,表明转染DCC基因功能区可以明显诱导SW1116细胞凋亡。
2.3 TUNEL法检测转染细胞凋亡情况 TUNEL法结果显示,实验组中可见大量被染成棕褐色的凋亡细胞,而质粒对照组和细胞对照组中基本没有,见图2。表明DCC基因功能区的表达可以明显诱导SW1116细胞凋亡。

图1 转染pIRES2-EGFP-DCC的SW1116细胞AO/EB染色荧光照片(×100)
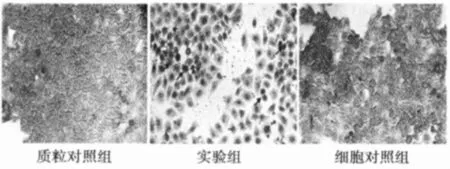
图2 转染p IRES2-EGFP-DCC的SW1116细胞TUNEL检测照片(×200)
3 讨论
DCC基因是Fearon等〔3〕在研究结直肠肿瘤时发现并命名的一个抑癌基因,编码由1 447个氨基酸组成的跨膜蛋白,其胞外区为神经生长因子-1(Netrin-1)受体,胞内区含有324个氨基酸,氨基端有25个疏水性氨基酸构成的信号序列,紧接着信号序列之后是4个免疫球蛋白样C2结构域,之后便是6个Ⅲ型纤连蛋白结构域。该基因定位于18号染色体长臂(18q21.3)。
研究发现结直肠癌组织中DCC基因缺失率在70%以上〔4〕;Gal等〔5〕在其研究中发现,对于 DCC 蛋白表达阳性的病人,当辅助于化疗时,可使生存率大大提高。将DCC反义RNA转染纤维母细胞,发现该细胞生长失控,体内注射对裸鼠具有致瘤性〔6〕。将DCC基因转染到结肠癌细胞,发现该细胞生长受到抑制,体内注射对裸鼠的致瘤性降低〔7,8〕。这些研究表明,DCC基因的表达可以明显抑制肿瘤细胞的增殖。
随后Mazelin等〔9〕发现,DCC对肿瘤的抑制作用是通过有条件诱导细胞凋亡实现的。Mehlen等〔10〕研究表明,DCC蛋白是一种依赖性受体,在配体Netrin-1存在时则具有抗凋亡作用,在配体缺乏时可以诱导凋亡。Mehlen等〔10〕进一步将DCC胞内区(DCC-IC)转染293T细胞,发现Caspases活性明显增加;体外Caspases孵育实验表明,DCC-IC在1 290 bp处被Caspases断裂;将断裂位点区域DCC-IC/1 121~1 290转染293T可以诱导细胞凋亡;进一步体外突变研究发现DCC-IC/1 243~1 264区段对于诱导凋亡是必需的。这些研究表明,DCC蛋白的肿瘤抑制作用是通过在配体Netrin-1缺乏时经Caspases通路诱导细胞凋亡实现的,而且其活性功能域很可能就是DCC-IC/1 243~1 264。DCC基因失活导致Bcl-2/Bax反常表达,抑制凋亡发生,可作为DCC基因凋亡分子机制之一〔11〕。
本研究通过脂质体法将含有DCC基因胞内区功能域(3 727~3 792 bp)的重组表达载体pIRES2-EGFP-DCC转染到结肠癌细胞SW1116,转染目的片段的实验组与对照组相比细胞增殖明显受到抑制;且实验组通过AO/EB法和TUNEL法分别检测到大量凋亡细胞,而对照组没有,证明该片段具有诱导肿瘤细胞凋亡的功能,为进一步研究DCC基因诱导细胞凋亡的机制打下了基础。
1 Lei T,Fang L,Cheng WQ,et al.Analysis of epidemiological characteristics and trends of colorectal cancer among elderly patients in ten cities and counties in China〔J〕.Disease Surveillance,2009;24(6):429-33.
2 Reale MA,Hu G,Zafar AL,et al.Expression and alternative splicing of the deleted in colorectal cancer(DCC)gene in normal and malignant tissues〔J〕.Cancer Res,2007;54:4493-501.
3 Fearon ER,Cho KR,Nigro JM,et al.Identification of a chromosome 18q gene that is altered in colorectal cancers〔J〕.Science,1990;247(4938):49-56.
4 Vogelstein B,Fearon ER,Hamilton SR,et al.Genetic alterations during colorectal-tumor development〔J〕.N Engl JMed,1988;319:525-32.
5 Gal R,Sadikov E,Sulkes J,et al.Deleted in colorectal cancer Protein expression as a possible Predictor of response to adjuvant chemotherapy in colorectal cancer patients〔J〕.Dis Colon Rectum,2004;47(7):1216-24.
6 Narayanan R,Lawlor KG,Schaapveld RQ,et al.Antisense RNA to the putative tumor-suppressor gene DCC transforms Rat-1 fibroblasts〔J〕.Oncogene,1992;12:1387.
7 Goyette MC,Cho K,Fasching CL,et al.Progression of colorectal cancer is associated with multiple tumor suppressor gene defects but inhibition of tumorigenicity is accomplished by correction of any single defect via chromosome transfer〔J〕.Mol Cell Biol,1992;12:1387-95.
8 Tanaka K,Oshimura M,Kikuchi R,et al.Suppression of tumorigenicity in human colon carcinoma cells by introduction of normal chromosome 5 or 18〔J〕.Nature,1991;349:340-2.
9 Mazelin L,Bernet A,Bonod-Bidaud C,et al.Netrin-1 controls colorectal tumorigenesis by regulating apoptosis〔J〕.Nature,2004;431(7004):80-4.
10 Mehlen P,Rabizadeh S,SniPas SJ,et al.The DCCgene product induces apoptosis by a mechanism requiring receptor proteolysis〔J〕.Nature,1998;395:801.
11 杨 莉,孙明军.DCC基因失活与大肠癌发病及与Bcl-2、Bax表达的相关性研究〔D〕.沈阳:中国医科大学硕士论文,2007.
