番茄抗病基因Ty-1的CAPS标记及检测
2012-05-22李海涛杜玉丽张子君邹庆道
李海涛 杜玉丽 张子君 曲 彤 邹庆道
(1辽宁省农业科学院蔬菜研究所,辽宁 沈阳 110161;2沈阳农业大学园艺学院,辽宁 沈阳 110161)
番茄(Lycopersicon esculentum Mill.)是一种世界性经济作物,品种多,产量高,营养丰富,用途广泛。限制番茄生产发展和产量的主要原因在于病害的流行为害和逆境条件的制约(叶青静 等,2009)。病毒病是番茄生产中最常见、发病最普遍的病害之一,其中的番茄黄化曲叶病毒病(Tomato yellow leaf curl virus disease,TYLCVD)是近年发生的一种暴发性、毁灭性病害,一旦发病,很难控制。番茄黄化曲叶病毒(Tomato yellow leaf curl virus,TYLCV)于1939~1940年在以色列首次发现(Pico et al.,1996),是一类由烟粉虱介导传播的双生病毒(geminiviruses),该病毒为双生病毒科(Geminniviridae)菜豆金色花叶病毒属(Begonurvints)成员(周雪平 等,2003)。番茄生长发育早期感染 TYLCV,其症状主要表现为植株生长缓慢或停滞,节间缩短,植株矮化,上部叶片变小、变厚、有褶皱、向上卷曲、变形、叶片边缘至叶脉区域黄化,无法正常开花结果,下部叶片症状不明显;后期感染TYLCV,染病植株仅上部叶片和新芽表现症状,坐果困难,果实僵化不膨大,或膨大速度极慢,果实变小,畸形果多,成熟期果实不能正常转色、失去商品价值,导致减产或绝收。目前在中东地区、非洲、亚洲、欧洲、美洲等世界各地都有发生并且已给美国、以色列、埃及、澳大利亚等国的番茄生产造成严重损失,我国广东、广西、台湾、上海、江苏等地都有发生的报道(何自福 等,2007;赵统敏 等,2007),并呈现由南向北迅速蔓延的趋势,已经成为影响番茄生产的主要限制因素之一。
目前番茄黄化曲叶病毒病的抗病基因有 Ty-1、Ty-2、Ty-3、Ty-3a、Ty-4、TY-5等,研究较多的是Ty-1、Ty-2、Ty-3基因。Zamir等(1994)认为Ty-1基因为主效基因,该基因为不完全显性单基因,并将其定位在番茄第6号染色体的RFLP标记TG297(4 cM)和TG97(8.6 cM)之间,图距6~10 cM。番茄黄化曲叶病毒病属检疫性病害,利用常规方法进行抗病育种有较大困难,利用分子标记结合常规方法能高效、准确地进行抗病材料的筛选与鉴定。本试验旨在寻找与番茄黄化曲叶病毒病抗病基因Ty-1紧密连锁的分子标记,并利用该标记进行抗病材料的检测,为分子标记辅助育种奠定基础。
1 材料与方法
1.1 材料
本试验共用番茄材料63份。其中含抗病基因Ty-1的纯合材料3份,编号为CK1、CK2、CK3,基因型为Ty-1/Ty-1,来自亚蔬中心(AVRDC);不含抗病基因Ty-1的感病纯合材料3份,编号为CK4(L402母本)、CK5(L402父本)、CK6(金棚1号母本),基因型为ty-1/ty-1,来自辽宁省农业科学院蔬菜研究所番茄课题组。
抗病的F18份,编号为7~14,来自上海种都种业科技有限公司,在2010年召开的“全国抗番茄黄化曲叶病毒病品种展示会”上表现为抗病。7号为迪达,8号为迪瑞,9号为TY4220,10号为迪维斯,11号为TY4378,12号为TY4430,13号为TY1415,14号为TY3175;未知抗病性的F125份,编号为 15~39,来自国外种子公司,具体名称不祥。未知抗病性的自交系材料24份,编号为40~63,来自辽宁省农业科学院蔬菜研究所番茄课题组。
1.2 方法
1.2.1 引物设计 番茄抗黄化曲叶病毒病抗病基因Ty-1的CAPS标记引物参照Zamir等(1994)的方法设计。引物由北京赛百盛生物技术公司合成。材料的特异性引物序列及其扩增片断见表1。
1.2.2 PCR扩增及酶切体系 DNA提取参照Williamson等(1994)的方法,由CTAB法提取。用于PCR反应和酶切的药品均购自tiangen生物工程公司。PCR反应总体积为20 μL,包括:模板DNA 20 ng,dNTP 0.4 μL,引物(10 U)各2.0 μL,Taq DNA聚合酶(2.5 U·μL-1)0.5 μL,Buffer 2.0μL,Mg2+2.0μL,终体积用超纯水加至20 μL。PCR的反应程序为:94 ℃预变性5 min;然后94 ℃变性1 min,64.6 ℃退火1 min,72 ℃延伸1 min,35个循环;最后72 ℃延伸10 min,扩增产物4 ℃保存。酶切体系为扩增产物7 μL加入10 U的Rsa I 酶1 μL,Buffer 2 μL,终体积用超纯水加至20 μL。37 ℃保温1.5 h。PCR产物及酶切产物于2.0%琼脂糖胶在电压5 V·cm-1条件下电泳30 min,终结果用Goodview 染色,Bio-RAD凝胶成像系统拍照。

表1 引物序列
1.2.3 CAPS标记的获得及抗病基因检测 利用设计的引物对3份抗病纯合材料CK1、CK2、CK3及3份感病纯合材料CK4、CK5、CK6进行PCR扩增,然后将扩增产物用Rsa I酶切,获得与抗病基因紧密连锁的CAPS标记;利用该标记分别对F1、自交系进行抗病基因的检测。
2 结果与分析
2.1 与Ty-1基因紧密连锁的CAPS标记片段的获得
6份材料的PCR扩增图谱如图1,酶切图谱如图2。
如图1显示,利用特异引物对6份抗病、感病材料进行PCR扩增,无论抗病材料还是感病材料均扩增出400 bp左右的片断,经RsaⅠ酶切后(图2),3份感病材料产生350 bp和50 bp左右的片段(但较小的50 bp的特异片段难以观察到,可能是目标条带产物相对较少,经酶切后小片段浓度较低所致)。3份抗病材料不能被 RsaⅠ酶切,仍呈现原来的 400 bp左右的片段。400 bp和350 bp两个片段能将抗病材料和感病材料加以区分,是与抗病基因连锁的标记。
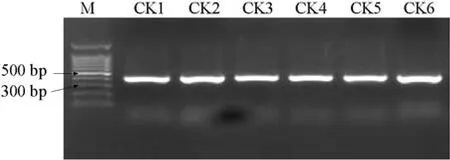
图1 6份材料的PCR扩增图谱
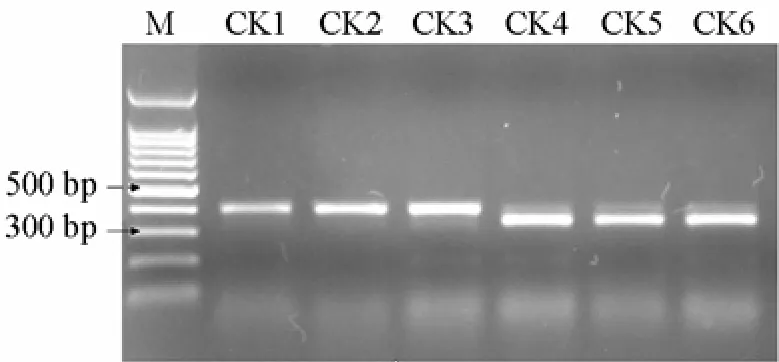
图2 PCR扩增产物的RsaⅠ酶切图谱
2.2 利用 CAPS标记对 8份抗病番茄 F1进行抗病基因检测
8份抗病材料的酶切图谱如图3。
如图3所示,8份抗病材料中,迪维斯、TY1415、TY31753份材料酶切后产生400、350 bp和50 bp左右的片段,说明这3份材料为含有抗病基因 Ty-1的杂交种;其他材料经酶切后只产生350 bp和50 bp左右的片段,和感病对照的扩增片段相同,说明这些材料不含抗病基因Ty-1。由于这些材料在田间均表现为抗病,说明这些材料可能含有除Ty-1以外的其他抗病基因。
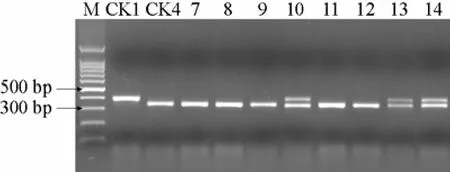
图3 8份抗病番茄F1的RsaⅠ酶切图谱
2.3 利用CAPS标记对25份未知抗病性的番茄F1进行抗病基因检测
25份材料酶切图谱如图4。
如图4所示,25份材料中有17份材料表现为杂合抗病型,酶切后产生400、350 bp和50 bp左右的片段,应是含有抗病基因Ty-1的抗病品种;8份材料表现为纯合感病基因型,酶切后产生350 bp和50 bp左右的片段,说明不含抗病基因Ty-1。
2.4 利用CAPS标记对24份番茄自交系进行抗病基因检测
24份材料酶切图谱如图5所示:24份材料均存在RsaⅠ酶切位点且酶切后均产生350 bp和50 bp左右的片段,与感病对照的扩增片段相同,说明这些自交系均不含抗病基因Ty-1。
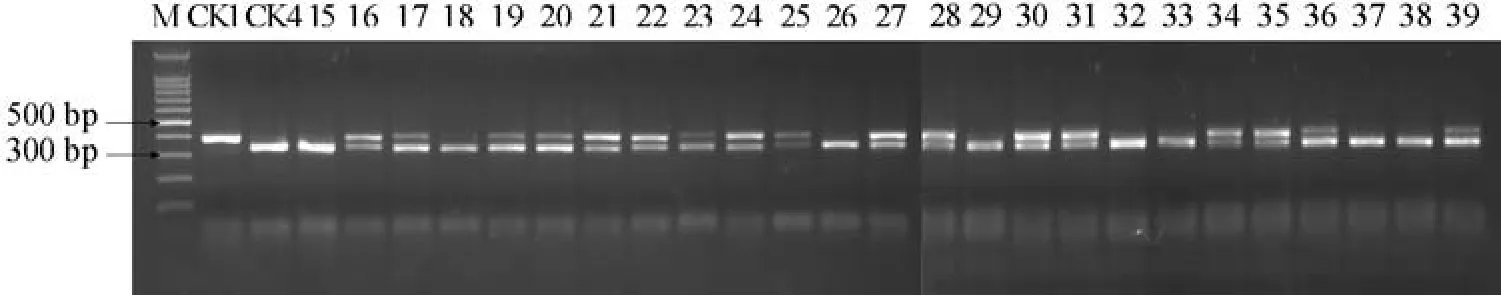
图4 25份番茄F1的RsaⅠ酶切图谱
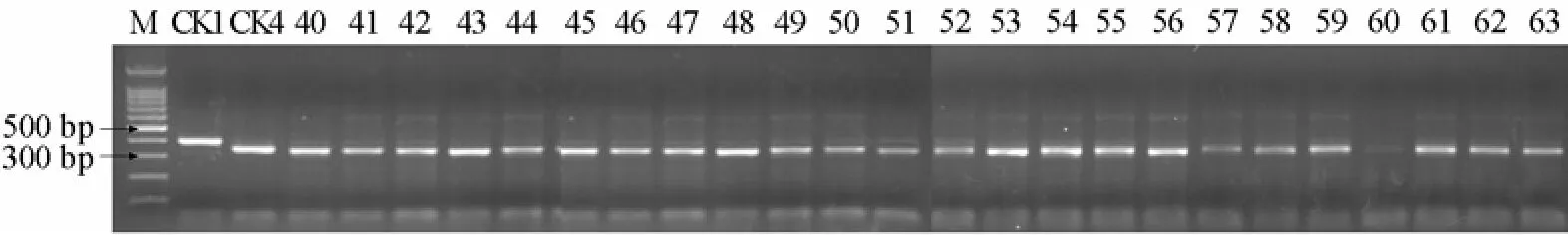
图5 24份番茄自交系的RsaⅠ酶切图谱
3 讨论
CAPS标记作为一种特异PCR标记,不仅比RFLP、AFLP等标记快速、简便,并且不用使用放射性同位素,减少了对人体的伤害,而且比RAPD、ISSR等PCR标记更稳定,特异性更强(于力 等,2008)。随着分子技术的迅速发展,利用分子标记辅助育种已极大地加快了育种的速度。
本试验找到了一个有效可用的 CAPS标记,为番茄黄化曲叶病毒病分子辅助育种奠定了基础,但该标记与抗病基因连锁距离的远近,尚需田间试验进一步验证。利用获得的标记共对33份番茄F1进行抗病基因的检测,有20份材料含有抗病基因Ty-1,13份材料不含Ty-1基因,含有 Ty-1基因的抗病品种多来自国外,说明国外抗病材料比较丰富。对 24份国内骨干育种材料进行检测,均不含抗病基因Ty-1,说明国内进行番茄抗TYLCVD育种不能直接利用以往的骨干自交系,需要进行材料创新。
不同地区的番茄黄化曲叶病毒的变异较大,导致同一个抗性材料在不同地区有不同抗性表现,甚至在同一地区不同年份间抗性表现不同。实践表明,选育抗病品种是防治TYLCV最为经济环保有效的方法。经过育种家多年的努力,国外在抗番茄黄化曲叶病毒病育种方面取得了显著成就。但是目前,抗TYLCVD的商业杂交种中导入单个抗性基因的居多,当病害大规模发生时,它们的抵抗能力依然下降。提高抗性水平的策略之一就是将多个抗性基因聚合到一个品种中,而有关专家已经证明了通过聚合不同抗源的基因可以提高番茄对TYLCV的抗性(Vdavski et al.,2008)。付蓉蓉等(2011)的研究已表明,同时含有纯合Ty-1和Ty-3抗性基因的番茄材料有更高更稳定的抗性。分子标记辅助选择的最大优越性表现在抗性基因累加系的构建中,将抗性基因累加到栽培品种是育种上培育持久抗性品种的有效手段之一(余文贵 等,2009)。并且随着番茄黄化曲叶病毒病的抗性基因 Ty-1、Ty-2、Ty-3、Ty-4及 Ty-5的定位及分子标记工作的进行(Zamir et al.,1994;Hanson et al.,2006;Ji et al.,2007,2009;Anbinder et al.,2009),育种学家可以利用相关的分子标记更加准确快速地筛选抗源材料,并结合传统育种将这些抗性基因聚合到一个品种中,从而培育出具有更高、更广、更持久抗性的番茄新品种(付蓉蓉 等,2011)。
付蓉蓉,刘杨,陈火英.2011.番茄黄化曲叶病的Ty-1和Ty-3抗性基因的PCR鉴定.分子植物育种(online),(9):1647-1652.
何自福,虞皓,毛明杰,罗方芳,林奕韩,王穗涛.2007.中国台湾番茄曲叶病毒侵染引起广东番茄黄化曲叶病.农业生物技术学报,15(1):119-123.
纠敏,周雪平,刘树生.2006.烟粉虱传播双生病毒研究进展.昆虫学报,49(3):513-520.
叶青静,杨悦俭,王荣青,李志邈,阮美颖,周国治,姚祝平.2009.番茄抗黄化曲叶病育种研究进展.中国农业科学,42(4):1230-1242.
于力,朱龙英,万延慧,杨少军,朱为民,薛林宝.2008.多重PCR技术鉴定番茄Ty-1和Mi基因.分子植物育种,6(20):165-169.
余文贵,赵统敏,杨玛丽,赵丽萍.2009.番茄黄化曲叶病及其抗病育种研究进展.江苏农业学报,25(4):925-930.
赵统敏,余文贵,周益军,季英华.2007.江苏省番茄黄化曲叶病毒病(TYLCV)的发生与诊断初报.江苏农业学报,23(6):654-655.
周雪平,崔晓峰,陶小容.2003.双生病毒—— 一类值得重视的植物病毒.植物病理学报,33(6):487-492.
Anbinder I,Reuveni M,Azari R,Paran I,Nahon S,Shlomo H,Chen L,Lapidot M,Levin I.2009.Molecular dissection of Tomato leaf curl virus resistance in tomato line TY172 derived from Solanum peruvianum.Theor Appl Genet,119(3):519-530.
Hanson P M,Green S K,Kuo G.2006.Ty-2 a gene on chromosome 11 conditioning geminivirus resistance in tomato.Tomato Genetic Cooperative Report,56:17-18.
Ji Y F,Schuster D J,Scott J W.2007.Ty-3,a begomovirus resistance locus near the Tomato yellow leaf curl virus resistance locus Ty-1 on chromosome 6 of tomato.Mol Breeding,20(3):271-284.
Ji Y F,Scott J W,Schuster D J,Maxwell D P.2009.Molecular mapping of Ty-4,a new Tomato yellow leaf curl virus resistance locus on chromosome 3 of tomato.Journal of the American Society for Horticultural Science,134(2):281-288.
Pico B,Diezm J,Nuez F.1996.Viral diseases causing the greatest economic losses to the tomato cropⅡ.The Tomato yellow leaf curl virus:a review.Sci Hortic(msterdam),67(3-4):151-196.
Vdavski F,Czosnek H,Gazit S.2008.Pyramiding of genes conferring resistance to Tomato yellow leaf curl virus from different wild tomato species.Plant Breeding,127:625-631.
Williamson V M,Ho J Y,Wu F F,Miller N,Kaloshian I.1994.A PCR-based marker tightly linked to the nematode resistance gene,Mi,in tomato.Theor Appl Genet,87:757-763.
Zamir D,Eksteinmi C,Heison I,Zakay Y.1994.Mapping and introgression of a Tomato yellow leaf curl virus tolerance gene,Ty-1.Theor Appl Genet,88(2):141-146.
