不同早籼稻品种幼苗期的耐冷生理鉴定
2012-05-09肖辉海郝小花吴小辉
肖辉海, 郝小花, 吴小辉, 侯 姣
不同早籼稻品种幼苗期的耐冷生理鉴定
肖辉海, 郝小花, 吴小辉, 侯 姣
(湖南文理学院 生命科学学院, 湖南 常德, 415000)
为了解4个早籼稻品种幼苗期耐冷性的生理特点, 在人工气候箱内经26 ℃(对照)、12 ℃、10℃和6℃处理4 d后, 测定分析水稻幼苗过氧化物酶(POD)活性、过氧化氢酶(CAT)活性、丙二醛(MDA)含量、抗坏血酸(AsA)含量、可溶性蛋白质含量及电解质渗漏率6个生理指标的变化及其与抗冷性的关系. 结果表明, 在相同低温胁迫条件下, 株两优120和金优L2两个早籼稻品种幼苗期POD活性、CAT活性、AsA含量及可溶性蛋白质含量均高于准两优143和湘早籼45号, 而MDA含量及电解质渗漏率均低于准两优143和湘早籼45号. 上述结果说明两系法杂交稻株两优120的耐冷性最强, 三系杂交水稻金优L2次之, 两系法杂交稻准两优143和常规水稻湘早籼45号耐冷性稍差.
早籼稻; 幼苗耐冷性; 生理指标; 人工气候箱
冷害是水稻生产中普遍发生的一种现象. 水稻苗期冷害是指: 在育秧期间, 遭到连续或短暂的低于秧苗正常生长发育温度影响, 使生长发育发生障碍甚至死亡的现象. 冷胁迫条件下, 水稻幼苗体内发生一系列适应低温的生理生化变化, 以增强其抗冷性和减小冷胁迫对其伤害[1-6]. 不同品种对低温的忍耐能力差异较大, 其中, 籼稻对低温的忍耐力明显弱于粳稻, 不同籼稻品种之间也有一定差异. 我国南方早春寒潮入侵, 气温骤然下降, 大部分早籼稻品种的幼苗不能适应这种变化无常的不良气候环境, 从而导致早籼稻幼苗的伤害和死亡. 特别在湖南, 早春寒潮入侵不仅频繁, 而且持续时间长, 更容易造成早籼稻幼苗严重冻害和死苗. 因此, 开展早籼稻苗期耐冷性的生理鉴定, 了解不同早籼稻品种幼苗对低温的耐受能力, 对于促进湖南水稻生产具有十分重要的现实意义. 本研究利用人工气候箱控温条件, 在准两优143、株两优120、金优L2和湘早籼45号4个早籼稻品种的幼苗期(3~4叶龄时)经低温处理后, 对幼苗组织中的过氧化物酶活性、过氧化氢酶活性、抗坏血酸含量、可溶性蛋白质含量、丙二醛含量及电解质渗漏率等进行测定, 并以此作为鉴定水稻苗期耐冷性生理指标, 对上述品种(组合)作苗期生理耐冷性探讨, 旨在为高产栽培上述品种(组合)提供指导.
1 材料与方法
1.1 材料
早籼稻品种(组合): 株两优120、金优L2、准两优143和湘早籼45号, 均由湖南亚华种业有限公司提供.
1.2 方法
1.2.1秧苗培育及低温处理
水稻种子用病害防治剂强六精(内含40%三氯异氰尿酸)500倍液浸种24 h, 30 ℃下催芽48 h, 5 d后将4个品种(组合)发芽一致(胚根1 cm, 胚芽0.5 cm)的稻种均匀播种在塑料盆中, 每塑料盆高10 cm、直径20 cm、容积1.75 L, 内装过筛稻田壤土1.25 kg. 每个品种(组合)播种40盆, 每盆播150粒, 在28 ℃(昼)/24 ℃(夜)(即日平均温度26 ℃)的人工气候箱中培养, 每天光照12 h(处理时为10 h), 光强8 000 lx, 相对湿度为75%~80%. 秧苗长至2叶1心后添加营养液, 每5 d换一次营养液. 当秧苗生长到3叶1心期, 将每个品种的秧苗分为4组进行温度处理: 一组在原温度(日平均温度26 ℃)条件下培养作对照, 其余的分别进行3个水平的冷胁迫处理, 即各自在12 ℃、10 ℃和6 ℃的冷胁迫下连续处理4 d, 然后取幼苗进行抗寒生理指标的测定.
1.2.2抗寒生理指标的测定
过氧化物酶(POD)活性的测定采用愈创木酚法,过氧化氢酶(CAT)活性的测定采用高锰酸钾滴定法, 电解质渗透率的测定采用DDS-307型电导仪法, 丙二醛(MDA)含量用硫代巴妥酸(TBA)法测定. 抗坏血酸(AsA)含量用2, 6-二氯酚靛酚滴定法测定. 可溶性蛋白质含量的测定采用考马斯亮蓝G-250法. 具体的操作都参照郑炳松[7]的方法进行.
水稻幼苗各项生理生化指标所反映的变化情况以6 ℃、10 ℃、12 ℃测定值对26 ℃测定值的相对变化率来表示, 即: 相对变化率(%)= 6 ℃(或10 ℃或12 ℃)测定值-26 ℃测定值/26 ℃测定值×100%.
2 结果与分析
2.1 冷胁迫对早籼稻幼苗POD、CAT酶活性的影响
过氧化物酶(POD)和过氧化氢酶(CAT)是水稻幼苗体内清除氧自由基、降低活性氧伤害的2种关键性酶, 它们的活性越高, 清除氧自由基的能力就越强, 对水稻幼苗的保护作用就越大, 反之则越小.
由表1可知, 在低温胁迫下, 4个早籼稻品种(组合)幼苗的POD活性的变化趋势基本一致, 即12 ℃和10 ℃下的POD活性都比26 ℃下的高, 其中以12 ℃下的POD活性最高; 而6 ℃下的POD活性都比26 ℃下的低(株两优120除外). 但4个早籼稻品种(组合)幼苗在低温胁迫下的POD活性与各自在26℃下的POD活性的相对变化率相差较大. 与26 ℃下的POD活性相比较, 株两优120幼苗在12 ℃处理后的POD活性的相对变化值最大, 分别比金优L2、准两优143和湘早籼45号高68.27%、100.06%和130.93%, 其次是金优L2和准两优143, 湘早籼45号的相对变化值最小. 10 ℃处理后的POD活性的变化情况与12 ℃处理后的的基本一致, 即POD活性由大到小的顺序是株两优120>金优L2>准两优143>湘早籼45号. 6 ℃处理后, 金优L2、准两优143和湘早籼45号幼苗的POD活性都比26 ℃下的低, 但下降幅度最大的是湘早籼45号, 其次是准两优143, 金优L2下降幅度最小. 而株两优120则仍然比26 ℃下的高.
由表2可知, 在12 ℃、10 ℃和6 ℃条件下, 4个早籼稻品种(组合)幼苗的CAT活性都比各自在26 ℃下的高, 但相对变化率相差较大, 其中, 株两优120在3种低温处理条件下的CAT活性相对变化值最大, 金优L2和准两优143居中, 湘早籼45号的CAT活性的相对变化值最小.
上述结果表明, 4个早籼稻品种(组合)在12 ℃、10 ℃和6 ℃ 3种低温胁迫下, 株两优120幼苗体内均保持相对高的POD、CAT活性, 金优L2和准两优143次之, 而湘早籼45号幼苗体内的POD和CAT活性相对较低.

表1 低温胁迫下早籼稻幼苗POD活性的变化
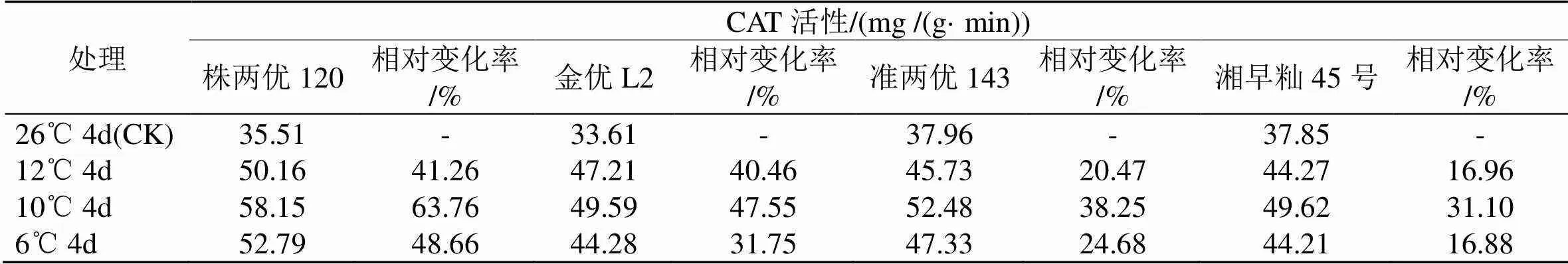
表2 低温胁迫下早籼稻幼苗CAT活性
2.2 冷胁迫对早籼稻幼苗丙二醛含量和电解质渗漏率的影响
植物在冷胁迫下的普遍反应是自由基清除系统物质(如POD、CAT等)的含量或活性下降, 膜脂过氧化产物(如丙二醛)含量上升, 电解质渗漏率增加. 因此, 丙二醛(MDA)含量和电解质渗漏率的变化可以反映植物遭受冷胁迫伤害的程度. 由表2可知, 在12 ℃、10 ℃和6 ℃条件下, 4个早籼稻品种(组合)幼苗的MDA含量和电解质渗漏率的变化趋势基本一致, 即随处理温度降低, 幼苗体内的MDA含量增加, 组织电解质渗漏率增大. 但4个早籼稻品种(组合)幼苗在相同低温处理下的MDA含量和电解质渗漏率与各自在26℃下的相比, 增加程度很不一致. 与各自在26℃下的MDA含量和电解质渗漏率相比较, 在相同低温处理下, 湘早籼45号的MDA含量和电解质渗漏率的相对变化值最大, 其次是准两优143, 再次是金优L2, 而株两优120的MDA含量和电解质渗漏率的相对变化值最小. 综上所述, 在冷胁迫下, 4个早籼稻品种(组合)幼苗体内的MDA含量和电解质渗漏率大小是: 湘早籼45号>准两优143>金优L2>株两优120. 说明在相同冷胁迫条件下, 湘早籼45号幼苗遭受冷胁迫伤害的程度最大, 准两优143和金优L2次之, 株两优120最小.

表3 低温胁迫下早籼稻幼苗中丙二醛含量的变化
2.3 冷胁迫对早籼稻幼苗抗坏血酸、可溶性蛋白质含量的影响
冷胁迫伤害的主要特征是低温对植物体内蛋白质的破坏. 在冷胁迫条件下, 植物细胞产生自溶水解酶, 加速蛋白质的分解过程, 造成蛋白质缺乏. 而抗坏血酸是植物体内普遍存在的一种抗氧化剂, 可降低或缓解植物因低温造成的伤害. 因此, 在冷胁迫下, 植物体内蛋白质、抗坏血酸含量的高低, 在一定程度上反映了植物的耐冷性的大小.
由表5可知, 4个早籼稻品种(组合)幼苗体内的抗坏血酸含量与各自在26 ℃下的相比较, 经12 ℃处理后, 株两优120、金优L2和准两优143幼苗的抗坏血酸含量比对照的高, 其中以株两优120提高的幅度最大, 金优L2次之, 准两优143提高的幅度最小, 而湘早籼45号则比对照的低. 在10 ℃和6 ℃下, 4个早籼稻品种(组合)幼苗抗坏血酸含量都比各自在26 ℃下的低, 下降幅度由大到小的顺序是: 湘早籼45号>准两优143>金优L2>株两优120. 说明12 ℃下4个早籼稻品种(组合)中, 株两优120和金优L2(尤其是株两优120)幼苗受冷伤害的程度最小, 准两优143次之, 而湘早籼45号受幼苗冷伤害的程度最大.

表4 低温胁迫下早籼稻幼苗电解质渗漏率的变化
表6显示, 在12℃下, 株两优120和金优L2幼苗的可溶性蛋白质含量比各自在26 ℃下的高, 且株两优120提高的幅度大于金优L2, 准两优143和湘早籼45号幼苗的可溶性蛋白质含量都比各自在26 ℃下的低, 但准两优143下降的幅度小于湘早籼45号. 在10 ℃和6 ℃下, 4个早籼稻品种(组合)幼苗的可溶性蛋白质含量都比各自在26 ℃下的低, 下降幅度由小到大的顺序是: 株两优120>金优L2>准两优143>湘早籼45号.
综上所述, 在冷胁迫下, 在供试的4个早籼稻品种(组合)中, 株两优120幼苗体内保持相对高的抗坏血酸含量和可溶性蛋白质含量, 抗冷性较强, 金优L2和准两优143次之, 而湘早籼45号幼苗体内的抗坏血酸含量和可溶性蛋白质含量水平相对较低, 抗冷性相对较弱.

表5 低温胁迫下早籼稻幼苗抗坏血酸含量的变化
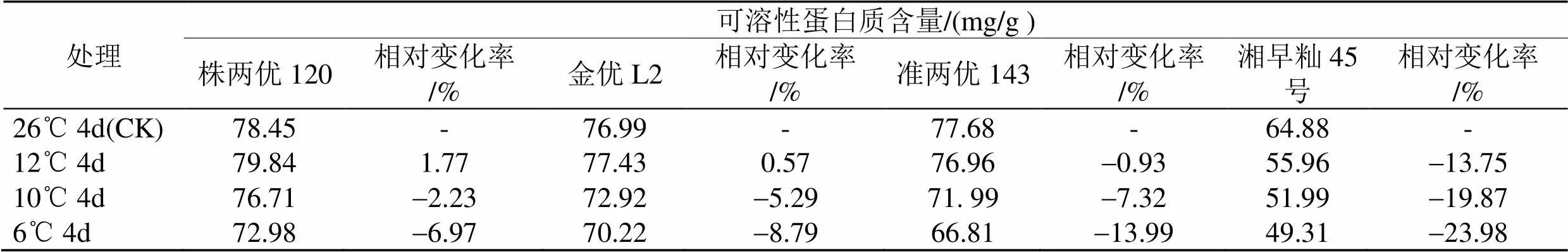
表6 低温胁迫下早籼稻幼苗可溶性蛋白质含量的变化
3 讨论
大量研究表明, 在正常情况下, 植物体内自由基的产生和清除之间处于平衡状态, 自由基水平很低, 不会伤害细胞. 但在低温冷害条件下, 自由基产生明显增加, 而自由基清除量下降, 导致自由基积累. 当自由基的积累超过域值时, 就会引起膜脂过氧化, 结果丙二醛 (MDA) 含量上升, 电解质渗漏率增加, 造成细胞酶保护系统的严重破坏[8-11]. 在水稻苗期如果遭遇冷害, 则导致秧苗失绿, 发僵, 分蘖减少, 秧苗枯萎甚至死苗等[12]. 因此, 在冷胁迫下, 水稻幼苗体内过氧化物酶(POD)、过氧化氢酶(CAT)、超氧化物歧化酶(SOD)等保护酶活性, 抗坏血酸(AsA)、还原型谷胱甘肽(GSH)等抗氧化剂含量, 以及MDA含量和电解质渗漏率等一系列生理生化指标的高低在一定程度上反映了水稻的抗冷性大小.
刘鸿先等[13]研究表明, 水稻幼苗中的SDS、POD、CAT活性随温度的降低而下降, 在耐冷性强的品种中其下降的程度小于耐冷性弱的品种. 同时GSH、AsA的含量也存在强耐冷品种的下降幅度小于耐冷性弱的品种的现象. 戴玉池等[14]研究发现, 在相同低温(6 ℃)胁迫条件下, 耐寒性强的品种金优402无论是电解质渗透率, 还是MDA的含量均低于耐寒性弱的品种新香优63和9910, 而可溶性糖含量、总叶绿素含量却明显高于新香优63和9910. 王晨光等[15]研究表明, 低温冷胁迫处理后, 抗寒性弱的品种黑粳7和龙盾94-652的POD活性显著低于抗寒性强的品种合江19, 且细胞膜透性增加, 细胞电解质渗透率大大增加. 傅泰露等[6]研究了5个不同类型水稻品种幼苗可溶性蛋白质含量与品种存活率(即耐冷性)之间的关系, 结果显示, 低温(8-10 ℃)胁迫下耐冷性较强的品种(如冈优527和冈优202)幼苗可溶性蛋白质含量高, 幼苗存活率也高; 反之, 耐冷性较弱的品种(如D优128和汕优63)幼苗可溶性蛋白质含量低, 幼苗存活率也低. 可溶性蛋白质含量相对值与品种的存活率(即耐冷性)呈极显著正相关.
水稻幼苗期耐冷性评价方法多以形态特征(如叶赤枯度)的主观评价和分级为主,准确性不高. 所以, 常通过测定与抗寒性相关的生理生化指标来进一步完善耐冷性鉴定体系,并通过分析抗寒相关生理生化指标和形态特征之间的关系来提高鉴定的准确性, 达到抗寒性鉴定的目的. 冷胁迫下水稻形态发生的变化, 源于其体内所发生的一系列生理上的变化, 因此, 本实验选用6个常用的抗性生理指标作为早籼稻幼苗期抗冷生理指标, 与实际相符. 从本实验采用的6个常见的抗寒生理指标的变化规律可见, 这6个指标皆呈规律性变化, 其中细胞电解质渗透率的变化幅度最大, 是反映早籼稻幼苗抗寒性的最灵敏的指标. 综合本实验6个生理指标可见, 在低温胁迫下, 4个供试早籼稻品种(组合)中, 两系法杂交早籼稻株两优120幼苗体内仍维持相对高的过氧化物酶和过氧化氢酶活性以及可溶性蛋白质和抗坏血酸含量, 而丙二醛含量和细胞电解质渗透率相对较低, 抗冷性最强, 其次为三系杂交早籼稻金优L2和两系法杂交早籼稻准两优143, 常规早籼稻湘早籼45号幼苗体内的POD活性、CAT活性、可溶性蛋白质含量及AsA含量最低, 而MDA含量及细胞电解质渗透率最高, 其抗冷性最弱.
[1] 刘鸿先, 曾韶西, 王以柔, 等. 低温对杂优水稻及其亲本幼苗中超氧物歧化酶的影响[J]. 植物学报, 1987, 29(3): 262-279.
[2] 曾韶西, 王以柔, 刘鸿先. 低温胁迫对水稻幼苗抗坏血酸含量的影响[J]. 植物生理学报, 1987, 13(4): 365-370.
[3] 王以柔, 曾韶西, 刘鸿先. 冷锻炼对水稻和黄瓜幼苗SOD、GR活性及GSH、AsA含量的影响[J]. 植物学报, 1995, 37(10): 776-780.
[4] 王晨光, 王希, 苍晶, 等. 低温胁迫对水稻幼苗抗冷性的影响[J]. 东北农业大学学报, 2004, 35(2): 205-207.
[5] 王国莉, 郭振飞. 低温对水稻不同耐冷品种幼苗光合速率和叶绿素荧光参数的影响[J]. 中国水稻科学, 2005, 19(4): 381-383
[6] 傅泰露,马均, 李敏, 等. 杂交水稻苗期耐冷性综合评价及其鉴定指标的筛选[J]. 西南农业学报, 2009, 22(3): 608-614.
[7] 郑炳松. 现代植物生生化研究技术[M]. 北京: 气象出版社, 2006.
[8] Chen S Y. Injury of merubrane lipid peroxidation to plant cell [J]. Plant Physiol Cummun,1991, 27(2): 84-90.
[9] Prasad TK. Mechanism of chilling-induced ox idative stress injury and tolerance: changes in anitioxidant system, oxidation of proteins and lipids and protease activities[J]. Plant, 1996, 10: 1017-1026.
[10] 彭昌操, 孙中海. 低温锻炼期间柑橘原生质体SOD和CAT酶活性的变化[J]. 华中农业大学学报, 2000, 19(4): 384-387.
[11] huang M, guo Z. Responses of antioxidative system to chilling stress in two rice cultivars differing in sensitivity[J]. Biologia Plantarum, 2005, 49(1): 81-84.
[12] 李霞, 程睿. 水稻不同生育期耐冷性鉴定[J]. 江苏农业科学, 2005(2): 23-26.
[13] 刘鸿先, 曾韶西, 王以柔, 等. 低温对杂优水稻及其亲本幼苗中超氧化物歧化酶的影响[J]. 植物学报, 1987, 29(3): 262-270.
[14] 戴玉池, 邓霞玲, 姜孝成, 等. 不同水稻品种幼苗期的耐寒生理鉴定及其利用[J]. 湖南师范大学自然科学学报, 2004, 27(3): 86-89.
[15] 王晨光, 王希, 苍晶, 等. 低温胁迫对水稻幼苗抗冷性的影响[J]. 东北农业大学学报, 2004, 35(2): 205-207.
Identification of chilling-tolerance physiology in different earlyrice seedings
XIAO Hui-hai , HAO Xiao-hua , WU xiao-hui , HOU jiao
(Department of Biology, Hunan University of Arts and Science, Changde 415000, China)
In order to understand the physiological characteristics of chilling tolerance of 4 early indica rice varieties at seedlingstage, changes of 6 physiological indicators [peroxidase (POD) activity, catalase(CAT) activity, ascorbic acid(AsA) content, soluble protein content, malondialdehyde(MDA) content and electrolyte leakage rate ] of rice seedlings and the relations of them to cold-resistance ability from 4 earlyrice varieties were studied 4 days after 26 ℃(CK)、12 ℃、10℃ and 6℃ treatment by using phytotron. The results show that the early indica rice varieties Zhuliangyou 120 and Jinyou L2 are obviously higher than the early indica rice varieties Zhunliangyou 143 and Xiangzaoxian 45 in POD activities, CAT activities, AsA contents and soluble protein contents under the same low temperature stress, but are obviously lower than the earlyrice varieties Zhunliangyou 143 and Xiangzaoxian 45 in the MDA contents and electrolyte leakage rates. The above results indicate that the two-line hybrid combination Zhuliangyou 120 had the greatest chilling tolerance and less in descending order three-line hybrid combination Jinyou L2, two-line hybrid combination Zhunliangyou 143 and conventional variety Xiangzaoxian 45.
earlyrice; chilling-tolerance of seedlings; physionogical indicators; phytotron
S 511
1672-6146(2012)01-0046-05
10.3969/j.issn.1672-6146.2012.01.013
2012-03-09
湖南省教育厅重点项目(09A066); 湖南省科技厅科技计划项目(2008FJ3066); 常德市博士创新计划重点项目(2010BS06)
肖辉海(1962-), 男, 博士, 教授, 主要从事植物发育生物学研究. E-mail: xxh-07@126.com
(责任编校: 谭长贵)
