复杂系统中的合作演化与自组织斑图
2012-03-22姜罗罗汪秉宏
姜罗罗, 汪秉宏
(1.温州大学物理与电子信息工程学院,温州 325035;2.中国科学技术大学近代物理系,合肥 230026;3.上海理工大学复杂系统科学研究中心,上海 200093)
复杂系统的研究具有天然的综合性和交叉性,既包括了物理学、生物学等自然科学学科,也包括了经济学、社会学等社会科学学科.统计物理为理解复杂系统中大量个体的集体行为提供了一些重要的概念和方法.在这些纷呈复杂的集体行为中,大量个体间的合作行为尤为引人瞩目.合作行为广泛存在于人类社会和动物世界,从单细胞的微生物到高级哺乳动物,从社会性昆虫到人类社会.个体为维持整体利益而付出部分自身代价的利他行为被称为合作;反之,个体为寻求自身利益的最大化而导致整体利益受损的行为被称为背叛.众所周知,合作是人类文明进步的基石.然而,根据达尔文的自然选择法则,自然界的生物个体趋向于采取能够最大化自身利益的背叛策略[1],这种背叛策略将导致整体利益受损.当体系中所有个体都选择背叛策略时,整体利益和个体利益都会受到最大程度的损害,这显然不能解释处处可见的合作行为.因此,理解合作行为成为复杂系统中最根本也是最具挑战性的课题之一[2].一些重要的问题就很自然地呈现在人们面前:自私个体间合作行为是如何自发涌现的;合作怎样演化;大量个体形成怎么样的空间自组织斑图等.
最近几年来,应用统计物理的概念和方法,人们在复杂系统研究领域内取得了令人瞩目的成就[3].对于复杂系统中合作与竞争现象的研究,人们通过对大量个体间的合作行为进行定量统计,研究其中所隐藏的统计性规律,并根据所研究的问题,提出基本假设,建立理论模型,来探索这些规律的产生机制和可能的动力学影响.基于这种统计物理的研究方法,一些重要的合作机制被发现,如群体选择(group selection)、亲缘选择(kin selection)、直接/间接互惠(direct/indirect reciprocity)、声望(reputation)、惩罚(punishment)等[4-6].特别是从2008年以来,仅在《Nature》、《Science》、《PNAS》、《PRL》等强影响因子期刊上就已经发表了10余篇文章[7-20].这些发现引发了人们从统计物理的角度进一步探索复杂系统中大量个体的合作演化及其自组织斑图.
1 演化博弈模型
von Neumann和Morgenstern在1944年出版的《博弈论和经济行为》一书为经典博弈论奠定了理论基础.他们提出了合作博弈,其基本思想就是指博弈双方的利益都有所增加,或者是一方得到的利益等于另一方失去的利益[21].合作博弈广泛存在于现实生活中,如签订劳动合同、商业契约等.我国古代的“田忌赛马”就是一个合作博弈的例子.《史记》记载,战国时期齐国大将田忌与齐国贵族赛马,设重金赌注.田忌听从兵法家孙膑的建议用下等马与贵族的上等马,上等马与贵族的中等马,中等马与贵族的下等马比赛.结果两胜一负,赢得重金.从博弈的角度看,在赛马中,田忌和贵族们商定3次赛马中赢的次数多的人获得赌金.在这个前提下,各人利用自己的马匹和策略进行比赛,结果田忌赢得的金钱等于贵族们失去的金钱.随后,Nash于20世纪50年代提出了非合作博弈,特别是Nash均衡的提出,使得经典博弈论有了非常优美的数学形式,并被广泛地应用于经济学领域.合作博弈强调群体理性(group rationality),就是从群体的角度考虑策略的选择,使得整体收益最大.所以合作博弈研究的是参与者在达成合作时如何分配合作得到的收益,即收益分配问题.而非合作博弈强调个体理性(individual rationality),就是从个体的角度考虑策略选择,使得个体收益最大.所以非合作博弈研究的是参与者在利益相互影响的情况下如何选择策略使自己的收益最大,即策略选择问题.在日常生活中,非合作博弈比合作博弈更为普遍,如价格战、军备竞赛、污染、贸易壁垒等.
尽管经典博弈论被广泛地应用于经济、政治等领域,但其在生态博弈中的不足之处是非常明显的.按照经典博弈论,生态系统中,雄性个体间争夺配偶的竞争将是以生命为代价的,因为它们被假定为超理性的,它们的策略不会更改.但现实中种群个体争夺配偶往往不会出现这种惨烈的情景.1973年,Smith等[22]提出了演化博弈,指出动物种群间的有限代价竞争能够促进物种整体的适应性.从此,演化博弈被广泛地用于研究各式各样的合作现象.本文将首先简要介绍社会复杂系统和生态复杂系统中演化博弈的模型,包括囚徒困境博弈、铲雪堆博弈、公共品博弈和石头-剪刀-布博弈.
1.1 囚徒困境博弈和铲雪堆博弈
囚徒困境博弈和铲雪堆博弈演化动力学可以通过以下的收益矩阵进行刻画.
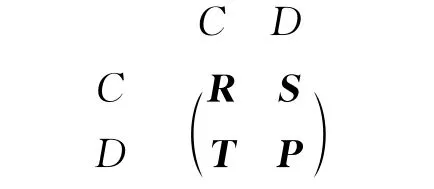
这里的矩阵元代表行方向上博弈参与者的收益,C表示合作者,D表示背叛者.两个合作者进行博弈后,两者均获得“奖励(reward)收益R”;两个背叛者进行博弈后,两者均获得“惩罚(punishment)收益P”;当合作者与背叛者进行博弈后,背叛者获得“诱惑(temptation)收益T”,而合作者则获得“被骗(sucker)收益S”.囚徒困境博弈与铲雪堆博弈的根本差别在于这些收益矩阵元大小的顺序不同.对于囚徒困境博弈,收益矩阵元的顺序为:T>R>P>S;而对于铲雪堆博弈,收益矩阵元的顺序为:T>R>S>P.
1.2 公共品博弈
假设有N个个体构成的体系拥有一个公共基金,每一个体可以选择投一个单位的货币量进入公共基金(合作),也可以选择不投资(背叛).如果有a个合作者,公共基金将在投资的基础上以r的倍数增值,即a·r个单位的货币.所得的货币由N个人平均分配.合作者之前付出了一个单位货币的投资,所以合作者获得的收益为(a·r/N)-1;而背叛者的收益为a·r/N.显然选择背叛策略的个体获得比合作更多的收益.但如果都选择背叛的话,大家的收益都为0.
1.3 石头-剪刀-布博弈
石头-剪刀-布博弈是非常简单的博弈形式,即3种策略循环抑制,其收益矩阵形式可以写为

1996年,美国加利福尼亚大学的行为遗传学家Sinervo和Lively首次发现真实生态系统中的石头-剪刀-布博弈[23].生活在美国加利福尼亚的雄性侧边斑点蜥蜴(side-blotched lizard)的喉部呈现出黄、橙、蓝3种不同的颜色,并且喉部不同颜色的雄性侧边斑点蜥蜴表现出不同的行为.黄喉雄性侧边斑点蜥蜴较为柔弱,行踪诡秘,在与其它亚种蜥蜴争夺配偶的竞争中扮演偷情者的角色;橙喉雄性侧边斑点蜥蜴较为生猛好斗,占据较大的领地,拥有为数众多的雌性配偶,奉行一夫多妻制;蓝喉雄性侧边斑点蜥蜴的武力介于其它两个亚种蜥蜴之间,同伴间相互合作捍卫自己的领地和雌性配偶,奉行一夫一妻制.在这个循环博弈中(见图1),橙喉雄性侧边斑点蜥蜴武力争夺蓝喉雄性侧边斑点蜥蜴的雌性配偶(石头抑制剪刀);蓝喉雄性侧边斑点蜥蜴武力捍卫自己的雌性配偶不受黄喉雄性侧边斑点蜥蜴袭扰(剪刀抑制布);由于橙喉雄性侧边斑点蜥蜴领地太大,顾此失彼,黄喉雄性侧边斑点蜥蜴得以偷偷摸摸地溜到没有设防的领地,找到雌蜥蜴成功交配(布抑制石头).这种雄性个体间繁殖策略的石头-剪刀-布博弈直接导致了各亚种的种群数量呈现周期振荡.Sinervo等发现,各亚种的种群数量在1990~1995年6年间呈现周期振荡.当蓝喉雄性侧边斑点蜥蜴的数量占据优势时,橙喉雄性侧边斑点蜥蜴的数量增加,从而抑制蓝喉雄性侧边斑点蜥蜴继续占据优势;随后橙喉雄性侧边斑点蜥蜴占据优势,黄喉雄性侧边斑点蜥蜴的数量增加,从而抑制橙喉雄性侧边斑点蜥蜴继续占据优势;紧接着黄喉雄性侧边斑点蜥蜴占据优势,蓝喉雄性侧边斑点蜥蜴数量增加,抑制黄喉雄性侧边斑点蜥蜴继续占据优势.如此往复,三亚种雄性蜥蜴互相制约,轮流居于优势地位,从而使蜥蜴总体数量上不会繁殖过快.尽管利他主义行为可能会损害某一只参加战斗的蓝喉雄性蜥蜴自身的繁殖机会,但是它却成全了其它蓝喉雄性蜥蜴与雌性交配,从而保护了它们的基因在下一代个体中能够继续存在[24].生态系统中石头-剪刀-布博弈的另一个经典范例是大肠杆菌3个亚种之间的循环制约竞争食物资源[25].

图1 三亚种雄性侧边斑点蜥蜴繁殖策略的石头-剪刀-布博弈示意图Fig.1 The rock-paper-scissors game in competitive reproduce strategies among three subspecies of Inner Coast Ranges lizards
2 空间演化博弈与自组织斑图
在演化博弈中个体如何更新自己的策略,是一个非常重要的研究内容.为了研究没有权威干预下的自私个体在什么条件下会合作,Axelrod[26]设计了以下收益矩阵的重复囚徒困境博弈.在两人进行博弈时,如果两人都选择合作策略,那么双方的收益都为3;如果两人都选择背叛策略,那么双方的收益均为1;如果一人选择合作而另一人选择背叛,那么选择合作策略的人收益为0,而选择背叛策略的人收益为5.Axelrod邀请其他科学家一起参加这个比赛,要求参加比赛的科学家把自己如何选择策略的想法编写成计算机程序,然后用单循环赛的方式将参赛程序两两博弈,以找出得分最高的策略选择规则.在博弈过程中,收益矩阵都是相同的,重复博弈的次数未知.第一轮比赛征集到了14个程序,每个程序运行300次.最终,加拿大学者Rapoport编写的“针锋相对(tit for tat,TFT)”策略选择规则获胜.“针锋相对”规则很简单,即第一次博弈选择合作策略之后选择对手上一次的策略.Axelrod等检查了程序运行结果后发现,“针锋相对”规则能够促进合作的产生.因为这个规则具有善良(niceness)、报复(retaliation)、宽容(forgiveness)以及清晰(clarity)的特点[26-27].个体一开始就选择合作,就是善良的体现,保持了合作的积极性,使得自身获得利益的同时也给对方利益.报复就是如果对手上一次选择背叛,那么下一次个体也会选择背叛,让对手明白是对方先错而付出的代价.宽容则有助于在对方背叛后重新开始合作,而简单清晰的规则易于被人理解,有利于长期合作.在两人静态囚徒困境中,使用“针锋相对”规则的个体其收益不可能超过对方,最多打个平手,但它的总收益最高.它赖以生存的基础是很牢固的,因为它让对方获得高收益.
在一个动态进化的群体中,这种合作能否产生、发展、生存下去;群体向合作的方向进化,还是向背叛的方向进化;如果大家开始都不合作,能否在进化过程中产生合作,对于这些疑问,Axelrod用生态学的原理来分析合作的进化过程.假设参与者所组成的策略选择规则是一代一代进化下去,进化的规则包括:第一,试错.人们在对待周围新环境时,起初不知道该怎么做,尝试不同的策略.第二,遗传.一个人如果合作性好,他后代的合作基因就多.第三,学习.在博弈过程中个体相互学习收益高的策略选择规则.如果“针锋相对”的策略选择规则收益高,那么就越多采取这种策略选择规则.按这样的思路,Axelrod设计了一个计算机实验,假设63个参与者中,谁在第一轮中的得分高,其在第二轮的群体中所占比例就越高,这就是该个体的适应性(fitness).这样,群体的结构就会在进化过程中改变,最终达到稳定.实验结果非常有趣,“针锋相对”起初在群体中仅占1/63,经过1 000代的进化结构稳定下来后,它占了24%[27].
Axelrod一方面揭示了“针锋相对”规则能够促使合作策略占据优势并稳定下来;另一方面,个体通过学习更新自己策略的思想使得人们更清晰地认识到合作机制研究的重要性,进一步激发了人们探索合作产生根源的激情.在真实世界中,个体并不能确定每一次跟同一个体进行博弈,同一时刻也可以同其他个体进行博弈,而且在真实世界中个体间的连接具有空间.因此,这样个体的策略选择需要考虑更多因素,譬如空间效应.无论在生态复杂系统中还是在社会复杂系统中,自私个体间的相互作用都使得体系出现自组织斑图,如合作团簇、螺旋波以及靶波.
2.1 社会复杂系统中的空间演化博弈
Nowak和May在1992年提出了忽略个体策略历史的囚徒困境博弈[28].个体分布在二维空间正方格子上,格点即代表个体,每一个体都有4个最近邻.每一轮博弈中,个体与最近邻进行囚徒困境博弈,并把该轮4次博弈所得的收益相加作为总收益.在下一轮博弈之前,个体对自己的总收益和近邻中最高总收益比较,如果近邻的最高总收益比自己的总收益大,那么个体的策略就跟随总收益最高的个体;若自己的总收益最高,则不改变自己的策略.这样N个参与者在二维空间规则正方格点上进行博弈.不失一般性,收益矩阵取为R=1,T=b(b>1),S=P=0.也就是说,两个合作者博弈时其收益均为1;合作者遇见背叛者时,合作者收益为0,背叛者收益为b;两个背叛者博弈时其收益均为0.这个模型只有一个参数b,用于刻画背叛者对合作者的优势.在N=L×L(L>20)无流边界的正方规则格子中,初始状态为合作者和背叛者随机分布.通过计算机模拟,Nowak和May发现,当b>1.8时,2×2或者更大的背叛者簇能够继续增大;而当b<1.8时,2×2或者更大的背叛者簇会减小.反之,当b<2.0时,2×2或者更大的合作者簇能够继续增大;而当b>2.0时,2×2或者更大的合作者簇会减小.在2.0>b>1.8的时候出现非常有趣的现象:合作者簇可以在被背叛者包围的情况下继续增长,而背叛者簇也能在被合作者包围的情况下继续增长.在这种情况下,由于合作簇和背叛簇的相互竞争,体系呈现出非常复杂的空间斑图,并且合作者所占比率最终在0.318附近.取中心有一个背叛者,其余均为合作者的情况为初始状态,体系呈现出具有对称结构的自组织斑图.图2(见下页)展示了这些空间斑图随时间t演化的情况,体系大小为99×99,无流边界条件,1.8<b<2.0.蓝色代表合作者坚持合作策略,红色代表背叛者坚持背叛策略,黄色代表合作者转变为背叛者,绿色代表背叛者转变为合作者.这些斑图都具有分形结构,随时间动态变化.在囚徒困境博弈中,由于个体仅限于与局域的近邻进行博弈,空间分布有利于合作.然而,这一结论并不对铲雪堆博弈有效[29].Hauert和Doebeli研究了具有空间结构和完全均匀混合(well-mixed)体系的铲雪堆博弈,发现当合作的付出-回报因子比较小的时候,空间效应有利于合作,即合作比率大于相同参数时完全均匀混合的合作比率;而当回报因子比较大的时候,空间效应抑制合作,即合作比率小于相同参数时完全均匀混合的合作比率.

图2 囚徒困境博弈中随时间演化的空间自组织斑图Fig.2 Evolution of spatial self-organized patterns in prisoners’dilemma games
基于Nowak等的工作,作者研究了空间囚徒困境博弈财富的不均匀性对合作的影响[30].个体分布在二维正方格点上,可以采取两个策略中的一种:合作(C)或者背叛(D).个体进行空间囚徒困境博弈的收益矩阵元仍为:R=1,T=b(b>1),S=P=0.个体i进行博弈获得收益Q后,其财富为Wi=Qα,Wj表示个体j的财富.此时,α因子代表着对收益的调控,如税收等.极端的例子为α=0,即所有个体最终的财富均为1.α=1时个体的收益没有被调控,α≫1则会出现贫富差距加大.进行策略更新的概率为

式中,K为噪音.
研究发现适当的α值能够促进合作,太大或者太小的α值都不利于合作.图3展示了α在较小值、中间值和较大值时,体系达到稳定状态时所呈现的自组织斑图.图中,L=100,b=1.01,绿色(淡色)代表合作者,蓝色(深色)代表背叛者.显然,合作者并不是孤立的分布,而是形成团簇.当收益没有被调节或调节太大时,即α=1.0和α=0.3(α为对收益的调控因子),体系中只有少量的合作者团簇;当α=0.5时,体系中出现很多合作者团簇,使得合作得以明显地促进.作者也研究了最大合作者簇的大小和合作者簇的平均大小随α的变化.尽管α太大或者太小时合作者簇消失,合作者簇的数目在很大范围内没有明显变化;然而,最大合作者簇的大小和合作者簇的平均大小随α变化非常明显,在α=0.5附近达到最大值.

图3 稳定状态时体系在不同α时的自组织斑图快照Fig.3 Typical snapshots of self-organized patterns at different values ofα
采用周期边界条件的二维规则正方格点代表参与博弈的个体,每一个体有4个近邻,共有N=L× L格点.每个格点被一个体或空位占据.空格代表空间资源可供个体迁移[31].空格密度d0定义为空位在所有格点中所占比率,那么人口密度则为(1-d0).采用随机方式异步更新个体策略状态的方法,每一次操作包含两个步骤:迁移和策略更新.在每一时间步内,平均所有个体都进行一次操作.随机选择一个体,计算其近邻中背叛者的数目nD用于确定其是否进行迁移.如果该个体近邻中有空位,该个体则以nD/4的概率向其近邻的空位迁移.如果该个体的近邻有多个空位,则随机选择一个空位迁移.若所有近邻都是空位,该个体将没有博弈的收益,则任意选择一个空位迁移,以期获得可能的博弈收益.迁移之后,个体以1-θ的概率学习其近邻中收益最高个体的策略.但如果其自身收益比其收益最高个体还高,则不学习,以θ(θ≪1)概率随机重新设置自己的策略.在这里θ扮演着噪声的作用,由环境噪声或者作决定时的不确定因素引起.值得注意的是,在策略更新中,受噪声影响和向更高收益个体学习是两个排他的过程,即个体以1-θ的概率学习而以θ的概率重新设置自己的策略.
自适应迁移能否使合作簇有效地抵御少数背叛者的入侵.为了回答这个问题,作者研究了特殊初始条件下的斑图演化.在这种特殊的初始条件中,一个包含大多数人口的合作簇中有极少量的背叛者.为了更好地评估自适应迁移的有效性,还对比了随机迁移的情况,如图4(a)(见下页)所示.在随机迁移的情况下,背叛者可以从合作簇的内部瓦解合作者之间的联系,而且随机迁移的合作者也不利于合作簇的形成.最终,背叛者占据整个系统.相反,对于自适应迁移,如图4(b)(见下页)所示,背叛者的数量仅仅是在刚开始的时候有所增加,但最终合作者把背叛者从合作簇中驱赶出去.在演化过程中,起初合作簇受到背叛者的冲击,合作簇被分割成小块;随后,由于个体迁移,合作者和背叛者被越来越多的空位隔离,背叛者从近邻中获得的收益也随之减少,所以背叛者变得不稳定;最终合作者占据绝对优势.对比图4(a)和4(b),可以得出这样的结论:自适应迁移能够促进合作,而随机迁移则有利于背叛.
图4(c)和图4(d)(见下页)解释了为何空位密度可以极大地影响体系的合作频率.特别地,在图4(c)中,背叛者的增加使得合作簇更加紧凑.但由于有限的空位,在大合作簇中的合作者难以迁移,所以没有足够的空位把背叛者和合作者隔离开来.尽管合作簇仍然能够抵御背叛者的入侵,但是,背叛者可以通过与其近邻的合作进行博弈而获得较高的收益.这样背叛者与合作者的收益就旗鼓相当,从而阻止了合作簇的进一步扩张.另一方面,当体系中有很多空位的时候,如图4(d)所示,合作者有很多可能的空位可以选择来迁移,这样合作者之间都很难接到,也就难以形成大的合作团簇.所以,大的空位密度抑制合作,而有利于背叛.
2.2 基于生态博弈的物种多样性与空间斑图
生物多样性是生态学的最核心问题之一.从真实生态系统中提炼出来的石头-剪刀-布博弈为研究生物多样性提供了一个非常重要的平台.假设一个体系中有A,B,C三物种,这3个物种形成相互循环捕食的生态链,这也类似于三物种循环捕食的Lotka-Volterra模型.当各物种间的相互作用(反应速率)是相同的时候,可以写出物种个体间博弈的收益矩阵.当系统演化达到平衡状态后,如果三物种仍然能够共存,那么就说明生物多样性得到了维持;反之,如果某一物种消失,那么说明生物多样性被破坏了.Reichenbach等研究了完全均匀混合(wellmixed)情况下A,B,C三物种进行石头-剪刀-布博弈的演化情况.他们发现,在三物种等量均匀分布的初始状态下,依据确定性的平均场方法物种不会出现灭绝,而随机过程的数值模拟得到的结果却是会出现物种灭绝.在石头-剪刀-布博弈中,相互循环抑制是维持物种多样性的重要机制.物种多样性的维持除了受噪声的涨落影响外,还受物种个体空间分布的影响.这种物种个体的分布,特别是自组织结构,对物种多样性的影响有待于进一步研究[32].
受大肠杆菌和阿米巴虫cAMP周期调制实验的启发,作者研究了受周期调制的石头-剪刀-布博弈中的自组织斑图结构,以揭示其特征和形成的原因[33].周期节律是体系中局部不均匀引起的,具有固定的频率.研究发现,这种局部的周期驱动可以使得全局出现非常规则的自组织结构——靶波.根据局部驱动频率与全局靶波率的关系,靶波的形成有3种模式:a.局部驱动频率与全局靶波频率同步;b.局部驱动频率与全局靶波频率间歇同步;c.局部驱动频率远大于全局靶波频率.研究表明可以通过调节驱动频率对斑图进行控制,同时也揭示了循环竞争物种间形成斑图选择的部分规律.
与Reichenbach[32]等的工作一样,作者也是在石头-剪刀-布博弈的框架下研究三物种的循环竞争.L×L的规则格子代表移动个体或空格.这些移动个体属于三物种中某一物种,三物种分别表示为A,B,C.空格则代表个体繁殖下一代所需的空间资源,表示为⊗,⊙表示任意物种或空格.个体间(或个体与空格)发生3个可能的过程:捕食、繁殖、迁移.捕食——物种A的个体以速率1捕食物种B的个体,那么B个体所在的位置就变为空格;同样方式,物种B的个体以速率1捕食物种C的个体;物种C的个体以速率1捕食物种A的个体.繁殖——个体以速率1在近邻的空格上产生一个后代.迁移——相邻的两个体或空格以速率γ交换位置.只有近邻间的个体(空格)才能发生这些相互作用的过程,并且可以用以下的反应式表达.
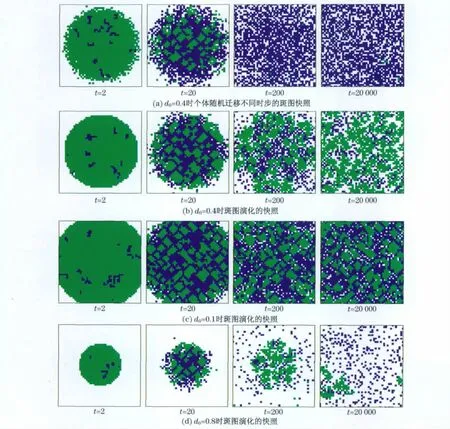
图4 不同空位密度情况下囚徒困境博弈中的空间斑图演化Fig.4 Evolution of spatial patterns for different values of vacation density in prisoners’dilemma games注:L×L=50×50.绿色(淡色)代表合作者;蓝色(深色)代表背叛者;白色代表空位.t表示系统演化的时间.

根据随机行走理论,个体的移动可以通过交换位置的速率进行定义,M=2γ/L2,表示移动个体在单位时间内走过的面积.与确定性过程不同,引入随机演化算法.在确定性算法下,演化时间被认为是连续的,系统不存在涨落(fluctuation).在随机算法中,瞬时过程类似于随机行走,系统存在涨落.常用的随机算法由Gillespie在1976年提出.该数值算法最初用于计算化学反应方程式,认为在分子间是随机发生化学反应的.在作者的数值算法中,捕食和繁殖均以1/(γ+2)的概率发生,而迁移(交换位置)则以γ/(γ+2)的概率发生.如果不考虑个体的分布不具有空间结构,而认为是完全均匀混合的(well-mixed),那么体系可以用偏微分方程组描述.以下的结果是在L×L规则正方格子上,无流边界,Gillespie随机算法,通过Monte Carlo数值模拟获得的.在每一Monte Carlo时间步内,首先随机选择一个体,然后随机选择其4个近邻中的一个体(空格),根据Gillespie算法确定是否发生捕食、繁殖、迁移或者什么也不发生.一个Monte Carlo时间步由N=L2这样的操作组成,平均每个格点都一次被随机选到.
在石头-剪刀-布博弈中引入局部不均匀的周期节律,通过在注入区周期注入三物种的个体,在小区域就形成一个周期节律对整个体系进行驱动.周期节律定义如下:在时刻t=0,注入区域被A物种的个体所占据;在时刻t=T0(T0表示注入物种的时间)注入区域被B物种的个体所占据;在时刻t=2T0,注入区域被C物种的个体所占据;在时刻t=3T0,注入区域又被A物种的个体所占据;如此往复,注入周期T=3T0.在n T0<t<m T0时间间隔内,其中m=n+1(m和n为大于0的自然数),注入区的个体仍然由Monte Carlo数值模拟更新.作者研究了不同的M和T0值的情况下系统的自组织结构.值得注意的是,当没有周期驱动的时候,模型得到的结果与Reichenbac等人的结果是一致的,即在迁移率M小于临界值时,物种多样性能够得到维持;而M大于临界值时,物种多样性被破坏.
如图5(a)所示,在参数M=10-4(M=2γ/L2表征个体的迁移快慢),T0=150和L=500下,随机的初始状态在周期节律的驱动下,逐渐被高度有序的靶波所代替(t=600T0).图中,黄色、红色和蓝色分别代表物种A,B和C;灰色代表空格.在演化过程中,t=T0时,位于体系中心的注入区开始注入物种A;t=2T0时,注入物种B;t=3T0,注入物种C;t=4T0,注入物种A;如此往复,注入区开始出现靶波.当t=200T0时,注入区的靶波开始向外扩大,但由于外围仍然是三物种的个体比较无序的分布靶波被约束在很小的范围内.当t=400T0时,靶波进一步扩大,最终在t=600T0时,靶波占据整个空间.从图5(b)中靶波形成的时空斑图,可以获得靶波形成的清晰图景.为了清晰起见,t1≤t≤t2是不连续的,t1和t2分别对应于33T0和466T0.在t<t1时间内,时空斑图比较紊乱,表明这一时期体系还没有出现规则的自组织结构;在t1≤t<t2时间内,中心区域的规则自组织结构开始向外扩大;当t≥t2,体系出现非常规则的自组织机构,这种结构随着时间演化,由里到外传播.当体系出现靶波之后,三物种所占比率随时间的变化呈现周期振荡,如图5(a)所示.与此同时,注入区三物种所占比率随时间变化也呈现周期振荡.这一周期振荡的频率与周期节律的频率相同.图5(b)是在靶波占据整个体系后物种A在整个体系(全局)和注入区(局部)所占比率随时间的振荡图.研究发现,靶波占据整个体系后,物种A的全局振荡与局部振荡同步,其它物种所占比率也有同样的结果.

图5 靶波的涌现过程Fig.5 Emerging of target waves
物种多样性和生物个体的空间分布是生态体系的两个主要特征.最近,种群的空间分布不均匀受到越来越多的关注,因为它与物种演化的稳定性和多物种共存有着密切联系.两个重要的因素影响着生态体系的生物多样性和物种的空间分布.第一个因素是物种间的捕食强度(相互作用强度).在生物链中,不同物种具有不同捕食强度.Huntley和Kowaleski研究了海洋动物显生宙地质年代的化石记录,发现捕食(竞争)强度与生物多样性具有强关联.最近的理论研究也聚焦于不同竞争强度的情况下,从物种多样性向物种单一性转化的相变是如何发生的,以及物种个体的空间自组织结构如何演化的问题.第二个因素是噪声.噪声能够影响物种个体空间自组织结构的形成,进而影响物种多样性的维持.为了研究竞争强度对物种多样性和自组织斑图的影响[34],作者修改了此前的反应式:个体以p/(p+q+γ)的概率竞争,以q/(p+q+γ)的概率繁殖,以γ/(p+q+γ)的概率迁移,p、q、γ分别代表发生竞争、繁殖和迁移的速率,如式(4)~(6)所示.
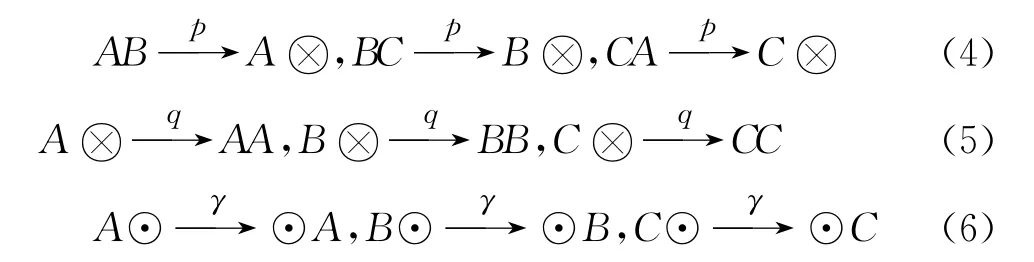
除了以上的随机方法外,基于扩散的偏微分方程也可以描绘以上过程.

式中,ρ0表示空位所占比率,D表示扩散系数,z和t分别表示个体所处的位置和演化时间.
图6展示了不同竞争强度p值的情况下体系演化到稳定状态后的自组织斑图.图6(a)为随机方法模拟所得,图6(b)为解偏微分方程所得.当p=0.1和1.0时体系出现全局有序的螺旋波;而当p=10.0时全局有序螺旋波破碎成无序的小螺旋波.可以通过观察螺旋波的波长变化获得更深入的理解.螺旋波波长被定义为:λ=X/L,其中X表示两相同物种形成的波的前直线距离(见图6).通过大量数值模拟发现,螺旋波的波长随着p的增大而减小,同时螺旋波的边缘变得越来越粗糙,这两个因素最终导致了p较大时螺旋波破碎.当p>pc时,全局有序的螺旋波破碎成为无序的小螺旋波.有趣的是,在生态系统中的石头-剪刀-布博弈中,空间拓扑结构和物种间的疾病传播也对物种多样性和空间斑图有较大影响[35-38].
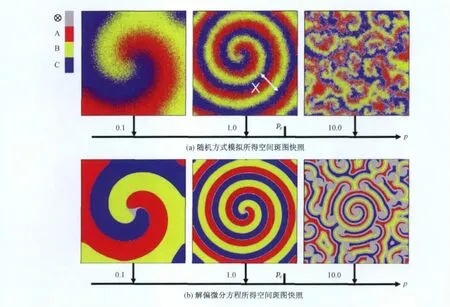
图6 不同竞争强度值下的空间斑图快照Fig.6 Typical spatial patterns emerging for different competition rates
3 复杂网络上的合作演化
由于人类处于复杂的人际关系中,仅仅通过空间斑图来理解合作行为是不够的,人际关系可以用复杂网络加以描述.网络结构对合作行为的影响,在复杂网络方面,人们在小世界网络和异质网络上进行了深入研究,发现网络的拓扑结构极大地影响着系统中个体间的合作程度.兰州大学的吴枝喜和汪映海等研究了NW(Newman-Watts)小世界网络上的囚徒困境博弈[39],发现个体在小世界效应的驱动下更倾向于参与博弈.浙江大学郑大昉和郑波发现小世界网络上适当地随机断边重连可以促进合作[40].中科大的任杰和汪秉宏等利用在小世界网络上交叉换边的方法,发现适当地随机换边能够促进囚徒困境博弈中的合作[41].北京大学的陈小杰和王龙基于学习理论发现适当的收益期望能够促进小世界网络上的合作[42].兰州大学的陈勇等发现在两个耦合的NW小世界网络中合作呈现出不同的同步行为[43].南昌航空大学的邱天等研究了有向小世界网络上的合作现象[44].吴枝喜和汪映海等则比较了随机网络、规则网络和无标度网络上的合作演化情况,发现无标度网络相对于其它两种类型的网络更有利于合作[45].北京大学的伏锋和王龙等研究了可以调节异质性的小世界网络,发现适当异质的网络拓扑结构能够促进合作[46].陈小杰和王龙等则进一步研究了异质的无标度网络上不同初始状态对合作演化的影响,当初始状态中合作者占据度大的节点时,最终背叛者趋于消失[47].复旦大学的李翔和周水根等发现无标度网络和具有团簇结构的网络均能够促进合作[48].上海交大的荣智海和汪小帆等发现网络中心节点间的连接能抑制合作,而非中心节点与中心节点间的连接能够促进合作[49].中科大的杜文博等发现加权无标度网络上的边权对合作具有较大影响[50].中科大汪秉宏研究组的王文旭、杨涵新、张海峰、史冬梅、刘润然和贾春晓也对复杂网络上合作演化动力学进行了深入研究[51-56].
与此同时,网络拓扑结构与博弈动力学共同演化也是人们关注的热点.北师大的李伟和胡岗发现当个体在博弈过程中连接局域收益最高的个体,能够让整个体系的拓扑结构呈现出自组织的无标度特性[57].北京大学的伏锋等发现个体选择学习对象以及博弈对象都对合作有较大影响[58-61].兰州大学汪映海、吴枝喜、黄子罡和关剑月等发现博弈动力学和偏好选择共同演化有利于合作[62-63].南京航空航天大学朱陈平等基于竞争个体形成无标度拓扑结构[64].厦门大学的杨冬平和帅建伟等研究了网络结构与博弈演化动力学共同演化机制,发现合作者的核心团簇对合作起关键的促进作用[65].
尽管网络结构和博弈动力学有了比较深入的探索,合作的稳定性仍然没有得到很好的理解.在具有平均度为4以及幂指数为-3的幂率度分布无标度网络中,基于公共品博弈理论,作者研究了合作在系统的连边受到攻击时的稳定性[66].在博弈动力学开始之前,先删除mL条与最大度节点相连的边.根据Albert等的工作,这种删除边的方式对应于网络受到攻击的情形.作者感兴趣的是删边数目如何影响合作的稳定性.在通常的网络动力学中,节点或者连边的毁坏将导致功能的级联失效.有趣的是,在公共品博弈动力学中,合作频率ρc先随删边数目的增加而减少,随后又随着删边数目的增加而增加[66].图7(a)(见下页)展示了在不同删边数目情况下体系中合作频率随收益增值因子r的变化情况,对于相同的r值合作频率在mL=0时具有最大值.从图7(b)(见下页)中可以更清晰地发现合作频率在r=1.8和r=2.0时先随mL增大而减少,然后随mL增大而增大.可见网络拓扑结构的毁坏导致合作的不稳定.
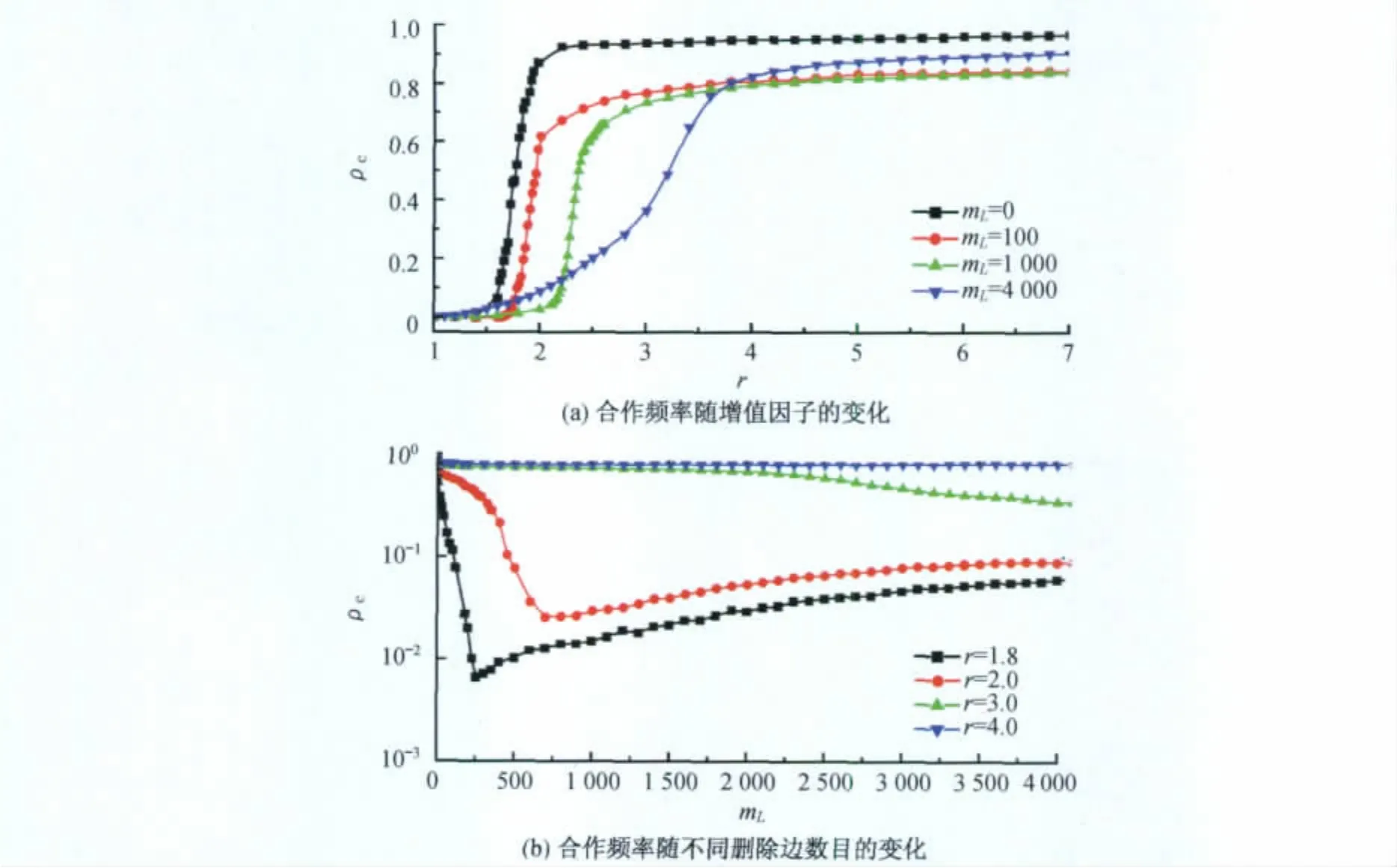
图7 合作频率随增值因子及不同随删除边数目的变化Fig.7 Fraction of cooperators varies with the multipli-cation factor and the number of deleted links
4 结 论
对于合作现象的研究,一方面合作机制仍在进一步的探索中;另一方面,一部分学者开始关注合作行为的传播及其稳定性.研究合作行为的传播,能够更深层次地理解合作机制.特别是目前发现的群体选择、亲缘选择、声望、惩罚以及空间效应等合作机制中究竟何种合作机制起主要作用并不清楚.研究合作传播及其稳定性能够定量衡量各种机制对合作的促进作用.
[1] Dugatkin L A.Cooperation among animals[M].New York:Oxford University Press,1997.
[2] Nowak M A.Evolutionary dynamics:exploring the equations of life[M].Harvard:Harvard University Press,2006.
[3] Castellano C,Fortunato S,Loreto V.Statistical physics of social dynamics[J].Rev Mod Phys,2009,81(2):591-646.
[4] SzabóG,Fáth G.Evolutionary games on graphs[J].Phys Rep,2007,446(4/5/6):97-216.
[5] Nowak M A,Sigmund K.Evolution of indirect reciprocity[J].Nature,2005,437(7063):1291-1298.
[6] Nowak M A.Five rules for the evolution of cooperation[J].Science,2006,314(5805):1560-1563.
[7] Santos F C,Santos M D,Pacheco J M.Social diversity promotes the emergence of cooperation in public goods games[J].Nature,2008,454(7201):213-216.
[8] King-Casas B,Sharp C,Lomax-Bream L,et al.The rupture and repair of cooperation in borderline personality disorder[J].Science,2008,321(5890):806-810.
[9] Cesarini D,Dawes C T,Fowler J H,et al.Heritability of cooperative behavior in the trust game[J].PNAS,2008,105(10):3721-3726.
[10] Reichenbach T,Frey E.Instability of spatial patterns and its ambiguous impact on species diversity[J].Phys Rev Lett,2008,101(5):058102.
[11] Clutton-Brock T.Cooperation between non-kin in animal societies[J].Nature,2009,462(7269):51-57.
[12] Rand D G,Dreber A,Ellingsen T,et al.Positive interactions promote public cooperation[J].Science,2009,325(5945):1272-1275.
[13] Helbing D,Yu W J.The outbreak of cooperation among success-driven individuals under noisy conditions[J].PNAS,2009,106(10):3680-3685.
[14] Wu J J,Zhang B Y,Zhou Z X,et al.Costly punishment does not always increase cooperation[J].PNAS,2009,106(41):17448-17451.
[15] Hamilton M J,Burger O,DeLong J P,et al.Population stability,cooperation,and the invasibility of the human species[J].PNAS,2009,106(30):12255-12260.
[16] Traulsen A,Hauert C,De Silva H,et al.Exploration dynamics in evolutionary games[J].PNAS,2009,106(3):709-712.
[17] Galla T.Intrinsic noise in game dynamical learning[J].Phys Rev Lett,2009,103(19):198702.
[18] Segbroeck S V,Santos F C,Lenaerts T,et al.Reacting differently to adverse ties promotes cooperation in social networks[J].Phys Rev Lett,2009,102(5):058105.
[19] Traulsen A,Semmann D,Sommerfeld R D,et al.Human strategy updating in evolutionary games[J].PNAS,2010,107(7):2962-2966.
[20] Kümmerli R,Burton-Chellew M N,Ross-Gillespie A,et al.Resistance to extreme strategies,rather than prosocial preferences,can explain human cooperation in public goods games[J].PNAS,2010,107(22):10125-10130.
[21] von Neumann J,Morgenstern O.Theory of games and economic behavior[M].Princeton:Princeton University Press,1944.
[22] Smith J M,Price G R.The logic of animal conflict[J].Nature,1973,246(5427):15-18.
[23] Sinervo B,Livel C M.The rock-paper-scissors game and the evolution of alternative male strategies[J].Nature,1996,380(6571):240-243.
[24] Sinervo B,Chaine A,Clobert J,et al.Self-recognition,color signals,and cycles of greenbeard mutualism and altruism[J].PNAS,2006,103(19):7372-7377.
[25] Kerr B,Riley M A,Feldman M W,et al.Local dispersal promotes biodiversity in a real-life game of rock-paperscissors[J].Nature,2002,418(6894):171-174.
[26] Axelrod R.The evolution of cooperation[J].New York:Basic Books,1984.
[27] Axelrod R,Hamilton W D.The evolution of cooperation[J].Science,1981,211(4489):1390-1396.
[28] Nowak M A,May R M.Evolutionary games and spatial chaos[J].Nature,1992,359(6398):826-829.
[29] Hauert C,Doebeli M.Spatial structure often inhibits the evolution of cooperation in the snowdrift game[J].Nature,2004,428(6983):643-646.
[30] Jiang L L,Zhao M,Yang H X,et al.Reducing the heterogeneity of payoffs:an effective way to promote cooperation in the prisoner’s dilemma game[J].Phys Rev E,2009,80(3):031144.
[31] Jiang L L,Wang W X,Lai Y C,et al.Role of adaptive migration in promoting cooperation in spatial games[J].Phys Rev E,2010,81(3):036108,.
[32] Reichenbach T,Mobilia M,Frey E.Mobility promotes and jeopardizes biodiversity in rock-paper-scissors games[J].Nature,2007,448(7157):1046-1049.
[33] Jiang L L,Zhou T,Perc M,et al.Emergence of target waves in paced populations of cyclically competing species[J].New Journal of Physics,2009,11(10):103001.
[34] Jiang L L,Zhou Tao,Perc M,et al.Effects of competition on pattern formation in the rock-paperscissors game[J].Phys Rev E,2011,84(2):021912.
[35] Ying C Y,Hua D Y,Wang L Y.Phase transitions for a rock-scissors-paper model with long-range-directed interactions[J].Journal of Physics A,2007,40(17):4477.
[36] Zhang G Y,Chen Y,Qi W K,et al.Four-state rockpaper-scissors games in constrained Newman-Watts networks[J].Phys Rev E,2009,79(6):062901.
[37] Yang R,Wang W X,Lai Y C,et al.Role of intraspecific competition in the coexistence of mobile populations in spatially extended ecosystems[J].Chaos,2010,20(2):023113.
[38] Wang W X,Lai Y C,Grebogi C.Effect of epidemic spreading on species coexistence in spatial rock-paperscissors games[J].Phys Rev E,2010,81(4):046113.
[39] Wu Z X,Xu X J,Chen Y,et al.Spatial prisoner’s dilemma game with volunteering in Newman-Watts small-world networks[J].Phys Rev E,2005,71(3):037103.
[40] Zhong L X,Zheng D F,Zheng B,et al.Networking effects on cooperation in evolutionary snow-drift game[J].Europhys Lett,2006,76(4):724.
[41] Ren J,Wang W X,Qi F.Randomness enhances cooperation:a resonance-type phenomenon in evolutionary games[J].Phys Rev E,2007,75(4):045101.
[42] Chen X J,Wang L.Promotion of cooperation induced by appropriate payoff aspirations in a small-world networked game[J].Phys Rev E,2008,77(1):017103.
[43] Chen Y,Qin S M,Yu L C,et al.Emergence of synchronization induced by the interplay between two prisoner’s dilemma games with volunteering in smallworld networks[J].Phys Rev E,2008,77(3):032103.
[44] Qiu T,Hadzibeganovic T,Chen G,et al.Cooperation in the snowdrift game on directed small-world networks under self-questioning and noisy conditions[J].Computer Physics Communications,2010,181(12):2057-2062.
[45] Wu Z X,Guan J Y,Xu X J,et al.Evolutionary prisoner’s dilemma game on Barabási-Albert scalefree networks[J].Physica A,2007,379(2):672-680.
[46] Fu F,Liu L H,Wang L.Evolutionary prisoner’s dilemma on heterogeneous Newman-Watts smallworld network[J].Eur Phys J B,2007,56(4):367-372.
[47] Chen X J,Fu F,Wang L.Influence of different initial distributions on robust cooperation in scale-free networks:a comparative study[J].Phys Lett A,2008,372(8):1161-1167.
[48] Li X,Wu Y H,Rong Z H,et al.The prisoner’s dilemma in structured scale-free networks[J].Journal of Physics A:Mathematical Theoretical,2009,42(24):245002.
[49] Rong Z H,Li X,Wang X F.Roles of mixing patterns in cooperation on a scale-free networked game[J].Phys Rev E,2007,76(2):027101.
[50] Du W B,Zheng H R,Hu M B.Evolutionary prisoner’s dilemma game on weighted networks[J].Physica A,2008,387(14):3796-3800.
[51] Wang W X,Ren J,Chen G R,et al.Memory-based snowdrift game on networks[J].Phys Rev E,2006,74(5):056113.
[52] Yang H X,Wu Z X,Zhou C,et al.Effects of social diversity on the emergence of global consensus in opinion dynamics[J].Phys Rev E,2009,80(4):046108.
[53] Zhang H F,Liu R R,Wang Z,et al.Aspirationinduced reconnection in spatial public-goods game[J].Europhys Lett,2011,94(1):18006.
[54] Shi D M,Zhuang Y,Wang B H.Group diversity promotes cooperation in the spatial public goods game[J].EPL,2010,90(5):58003.
[55] Liu R R,Rong Z H,Jia C X,et al.Effects of diverse inertia on scale-free-networked prisoner’s dilemma games[J].EPL,2010,91(2):20002.
[56] Jia C X,Liu R R,Yang H X,et al.Effects of fluctuations on the evolution of cooperation in the prisoner’s dilemma game[J].EPL,2010,90(3):30001.
[57] Li W,Zhang X,Hu G.How scale-free networks and large-scale collective cooperation emerge in complex homogeneous social systems[J].Phys Rev E,2007,76(4):045102.
[58] Fu F,Chen X J,Liu L H,et al.Social dilemmas in an online social network:the structure and evolution of cooperation[J].Phys Lett A,2007,371(1/2):58-64.
[59] Fu F,Hauert C,Nowak M A,et al.Reputation-based partner choice promotes cooperation in social networks[J].Phys Rev E,2008,78(2):026117.
[60] Fu F,Wang L,Nowak M A,et al.Evolutionary dynamics on graphs:efficient method for weak selection[J].Phys Rev E,2009,79(4):046707.
[61] Fu F,Wu T,Wang L.Partner switching stabilizes cooperation in coevolutionary prisoner’s dilemma[J].Phys Rev E,2009,79(3):036101.
[62] Wu Z X,Wang Y H.Cooperation enhanced by the difference between interaction and learning neighborhoods for evolutionary spatial prisoner’s dilemma games[J].Phys Rev E,2007,75(4):041114.
[63] Wu Z X,Xu X J,Huang Z G,et al.Evolutionary prisoner’s dilemma game with dynamic preferential selection[J].Phys Rev E,2006,74(2):021107.
[64] Zhu C P,Zhou T,Yang H J,et al.The process of coevolutionary competitive exclusion:speciation,multifractality and power-laws in correlations[J].New J Phys,2008,10(2):023006.
[65] Yang D P,Lin H,Wu C X,et al.Effect of mortality selection on the emergence of cooperation with network dynamics[J].New J Phys,2009,11(7):073048.
[66] Jiang L L,Perc M,Wang W X,et al.Impact of link deletions on public cooperation in scale-free networks[J].EPL,2011,93(4):40001.
