羊草生物量和养分分配对养分和水分添加的响应
2012-03-13黄菊莹余海龙袁志友李凌浩
黄菊莹,徐 鹏,余海龙,袁志友,李凌浩
(1.宁夏大学新技术应用研究开发中心,宁夏 银川 750021; 2.辽宁省环境保护宣传教育中心,辽宁 沈阳 110033;3.宁夏大学资源环境学院,宁夏 银川 750021; 4.中国科学院植物研究所植被与环境变化国家重点实验室,北京 100093)
降水格局改变、大气氮沉降增加以及由此引起的磷有效性改变[1]是当前全球变化生态学研究领域的热点。这三者改变了土壤水分和养分有效性,因此会影响植物养分吸收、分配和回收转移等策略。内蒙古多伦草原位于我国东北部,是我国典型的农牧交错带之一。该草原植被属于温带典型草原,主要由克氏针茅(Stipakrylovii)、冰草(Agropyroncristatum)、羊草(Leymuschinensis)等组成[2]。区域土壤供氮能力低下,年降水量385.5 mm,年蒸发量1 748 mm,因此该县草原生态系统主要受降水和氮的限制[3-4],对全球气候变化的反应敏感。全球范围的降水格局改变和大气氮沉降增加,不仅可能影响该脆弱生态系统的植被生长,而且可能改变该区域生物地球化学循环。而大气氮沉降持续增加,则可能使该脆弱生态系统由受氮限制转变为受磷限制的系统,导致区域植被磷需求增加[5]。因此在内蒙古多伦草原生态系统,开展沿水分、氮和磷梯度植物生物量和养分分配的响应模式的研究,对预测全球变化对该区域植被结构和功能的潜在影响具有一定的实践意义。
羊草是多年生根茎禾草,营养繁殖能力很强,是我国内蒙古典型草原群落的建群种或优势种[6-7]。本研究通过设置氮肥、磷肥和控水的盆栽控制试验,研究羊草地上、地下生物量和养分的响应格局,探讨环境变化对羊草生物量和养分分配的影响,旨为深入理解温带草原生态系统结构和功能对全球气候变化的响应机理提供基础数据。
1 材料与方法
1.1研究区自然概况 试验地点设在内蒙古锡林郭勒盟多伦县中国科学院植物研究所多伦恢复生态学实验示范研究站。样地所在地多伦县十三里滩(115°50′~116°55′ E,41°46′~42°39′ N)属于典型温带半干旱季风气候区,年平均降水量385.5 mm,其中70%集中在6―8月。年平均气温1.9 ℃,最冷月1月平均气温为-17.8 ℃;最热月7月平均气温为18.8 ℃。
1.2试验设计 试验于2006年5月中旬至11月初进行。以多伦县十三里滩自然草地的羊草幼苗为研究材料,分别布置了施氮肥(0、0.5、1、2、4、8、16、32、64和128 g·m-210个水平)、施磷肥(水平同氮处理)和水分供给(11.5、12.7、14.3、16.4、19.1、22.9、 28.7、38.2、57.3和114.6 mL·cm-210个供水量水平)的盆栽控制试验。具体试验设计参考文献[8]。数据分析中,分别以T1、T2、T3、T4、T5、T6、T7、T8、T9、T10代表各处理。以上各处理均5次重复。
1.3材料与方法 分别于2006年8月初和10月底,用剪刀齐地面剪下羊草地上部分,装袋带回实验室烘干(65 ℃,48 h)并称量。同期收集盆内羊草的地下部分,用水清洗干净,烘箱中烘干(75 ℃,72 h)并称量。所有干样粉碎后,采用全自动凯氏定氮仪(Kjektec System 2300 Distilling Unit, FOSS Tecator AB,Hoganas,Sweden)测定干样全氮,钼锑抗比色法测定干样全磷。
1.4统计分析方法 采用分段回归拟合生物量和养分库的变化趋势[9]。数据点为平均值±标准误。
2 结果
2.1氮、磷和水分供给改变对生物量及其分配的影响 增加氮肥、磷肥和水分供给均提高了羊草地上、地下生物量(图1~3):低供给情况下,两月地上生物量和地下生物量均呈增加趋势;之后随氮、磷和水分供给量的增加,地上、地下生物量逐渐降低。8月,各处理低水平下地上、地下生物量差别不大,之后随供给量增大地上生物量逐渐超过地下生物量。施氮、施磷和水分控制试验平均地下生物量分别为地上生物量的81.8%、86.1%和82.9%;10月,地下生物量显著高于地上生物量,3个试验平均地下生物量分别为地上生物量的1.5、2.3和2.3倍。
2.2氮、磷和水分供给改变对养分及其分配的影响 适量施氮肥显著提高了羊草地上、地下养分库(图4):低供给情况下,养分库均呈线性增加趋势;但当氮供应量继续增加时,养分库显著降低。8月地上养分库高于地下养分库;而10月,地下养分库则高于地上养分库。8月地上氮库是同期地下氮库的2.6倍,地上磷库是同期地下磷库的1.8倍;10月,地上氮库仅为同期地下氮库的70.4%,地上磷库则不足地下磷库的一半(49.2%)。

图1 羊草生物量随施氮量增加的变化趋势Fig.1 Changes of biomass of Leymus chinensis with N application gradient

图2 羊草生物量随施磷量增加的变化趋势Fig.2 Changes of biomass of Leymus chinensis with P application gradient

图3 羊草生物量随水分供给增加的变化趋势Fig.3 Changes of biomass of Leymus chinensis with water irrigation gradient
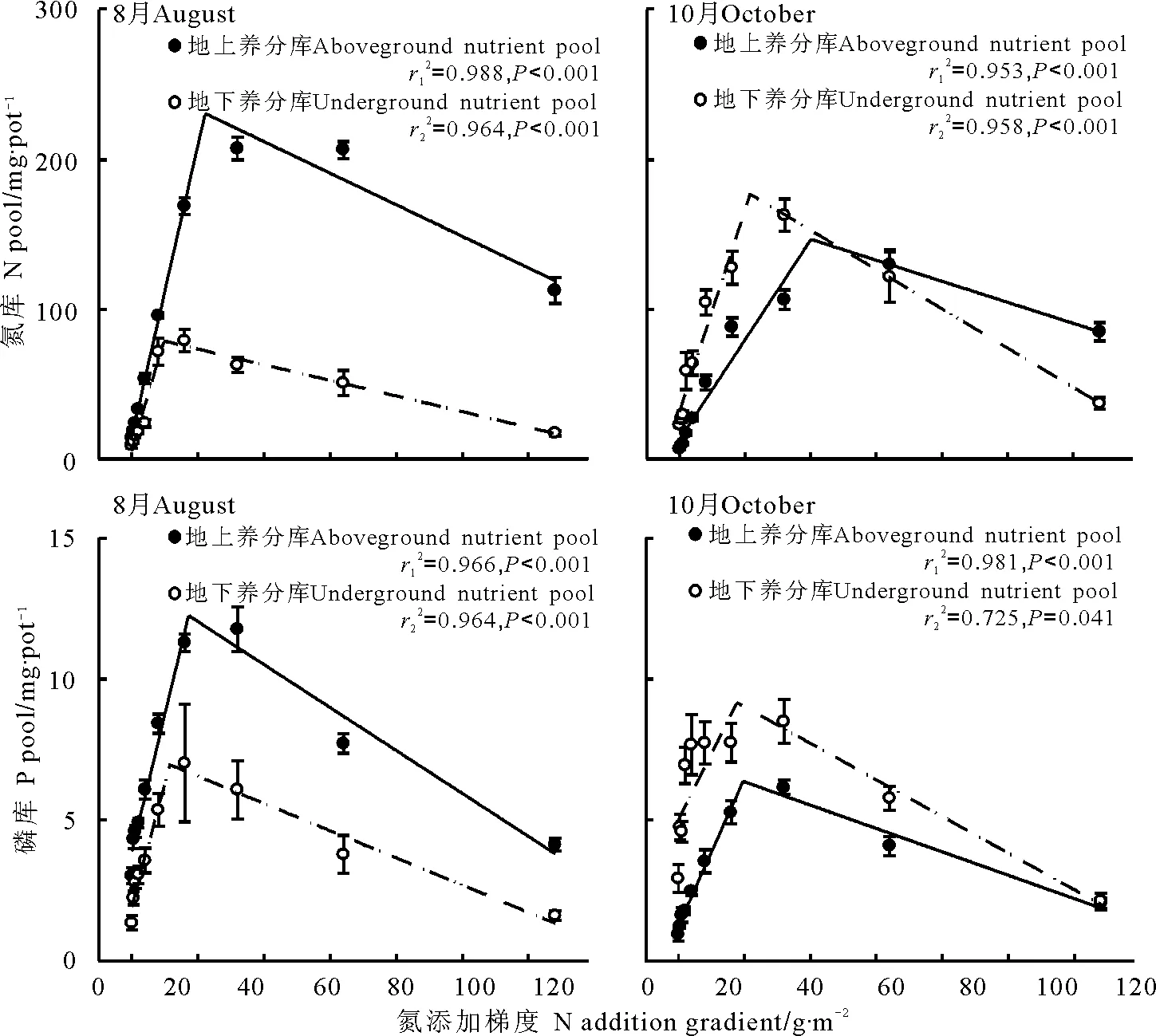
图4 羊草养分库随施氮量增加的变化趋势Fig.4 Changes of nutrient pool of Leymus chinensis with N application gradient
适量施磷肥显著提高了羊草地上、地下养分库(图5):低供给情况下,养分库均呈显著增加趋势;磷水平继续增加后,地上、地下养分库增加趋势变缓(磷库)或降低(氮库)。8月平均地上部分氮库和磷库分别为59.8和17.5 mg·盆-1,平均地下部分氮库和磷库分别为30.3和9.8 mg·盆-1,地上养分库高于地下养分库;10月地下磷库随施磷量增加逐渐超过地上磷库,平均地上部分氮库和磷库分别为27.1和21.6 mg·盆-1,平均地下部分氮库和磷库分别为66.8和21.5 mg·盆-1。

图5 羊草养分库随施磷量增加的变化趋势Fig.5 Changes of nutrient pool of Leymus chinensis with P application gradient
沿水分供给梯度,地上、地下养分库呈现先增加后降低或增加幅度变缓的趋势(图6):低供给情况下,养分库均呈增加趋势;随着供水量继续增加,8月地上、地下氮库呈降低趋势,地上、地下磷库增加幅度变缓。8月平均地上部分氮库和磷库分别为77.3和6.3 mg·盆-1,平均地下部分氮库和磷库分别为38.4和4.7 mg·盆-1,地上部分养分库高于地下部分;10月平均地上部分氮库和磷库分别为35.1和1.3 mg·盆-1,平均地下部分氮库和磷库分别为130.2和12.3 mg·盆-1,地下部分养分库高于地上部分。
2.3氮、磷和水分供给改变对根冠比的影响 氮、磷和水分添加改变了羊草根冠比(表1):施氮肥和施磷肥处理下,8月羊草根冠比较低,尤其是高氮和高磷处理;水分添加处理,8月份干旱胁迫下羊草具有较低的根冠比,10月份干旱胁迫下具有较高的根冠比。可见,施氮、施磷和水分添加处理对羊草根冠比的影响与季节有关。
3 讨论
一般而言,土壤养分(氮和磷)和水分有效性过高或过低均不利于植物生长[10-14]。本研究中,沿氮、磷施用梯度,羊草地上、地下生物量均呈先增加后降低的趋势(图1和图2),表明适量施氮、磷肥能显著促进羊草生长,但施用量过大会对羊草的生长发育产生抑制作用。生物量分配方面,在植物生长旺期(8月),一方面地上部分光合作用较高,使得干物质不断积累,另一方面地下部分不断输送养分给地上部分,因此促进了地上部分的生长;之后随着生长季的逐渐结束,植物地上部分开始枯死,有效光合面积减少,加之地上组织中的营养物质或淋失或转移给地下器官,促使地下部分生物量高于地上部分。本研究中,大体而言,8月地上生物量高于地下生物量,10月地下生物量超过地上生物量,说明施氮和磷肥降低了羊草根冠比,与其他研究结果一致[15-17]。研究结果表明,氮和磷限制着内蒙古温带草原生态系统植物生长。外源养分施用减轻或消除了氮和磷对植物生长的限制性,因此促进了羊草地上、地下部分生长;但当土壤养分有效性达到相对高的水平后,继续增加外源养分投入,则对羊草生长产生毒害,不利于各组织生长。控水试验中,沿水分梯度羊草地上、地下生物量呈先增加后降低的趋势(图3),说明适量增加供水减轻或消除了水分亏缺对植物生长的限制性,因此促进了植物生长[18-20]。而当土壤水分供给过多时(如水淹),植物的根系生长受到抑制[21-22],因此适宜的土壤水分可以促进植物生物量积累,而干旱胁迫则不利于植物生长[23-25]。干旱胁迫影响了植物生物量的分配:当植物处于干旱胁迫时,其生物量更多地分配给地下器官(如根系和根茎),导致根冠比增加[26-27]。本研究中,8月时11.5 mL·cm-2的供水量下,有明显高的根冠比(表1),表现出对极端干旱的高度适应性;但10月时根冠比随供水量的增加而增加(从1.75增加至2.70),这可能与冬季生物量由地上部分转移至地下部分有关。

图6 羊草养分库随水分供给增加的变化趋势Fig.6 Changes of nutrient pool of Leymus chinensis with water irrigation gradient
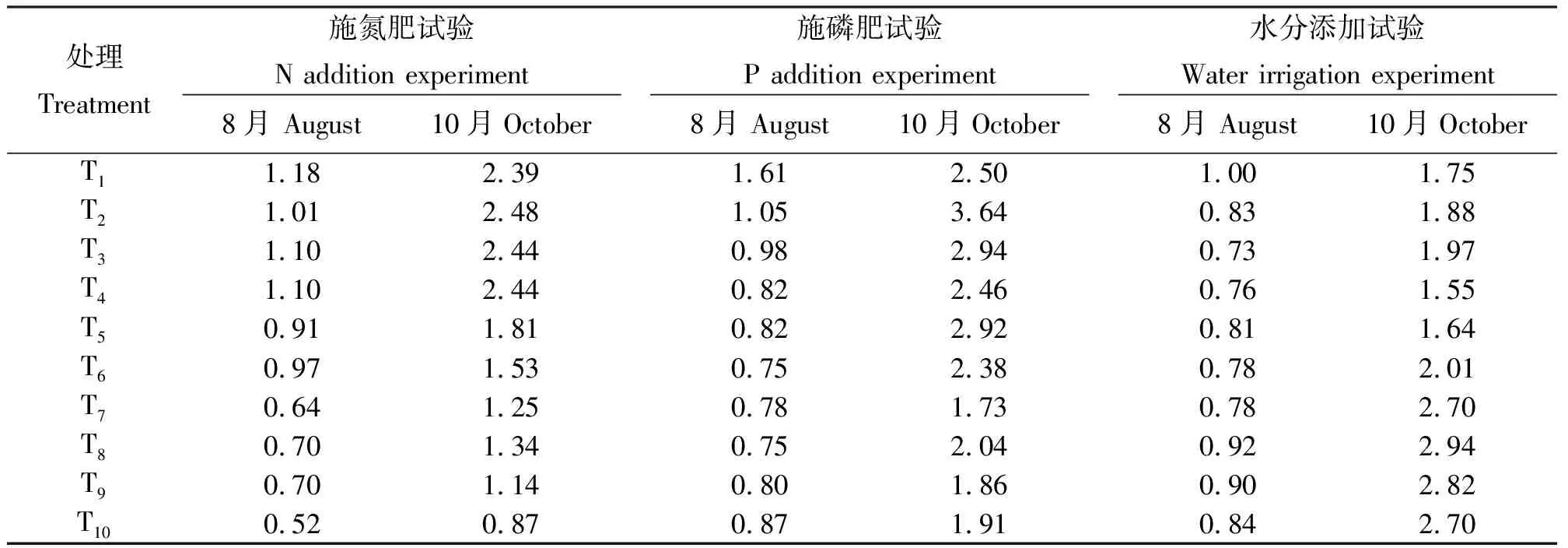
表1 氮添加、磷添加和水分添加对羊草根冠比的影响Table 1 Effects of N, P and water application on root to shoot ratio of Leymus chinensis
对多年生草本植物而言,在其生长季结束之前,地上部分枯死并将其内部养分转移给地下部分,达到养分保存的目的[28]。本研究中适量氮、磷和水分供给水平下,羊草地上、地下部分养分库均随氮、磷和水分供给量增加而增加。表明低养分和水分供给促进了8月地上、地下部分氮和磷的积累,增加了10月地上、地下部分归还给土壤的氮和磷含量。在养分分配方面,8月地上部分养分库高于地下部分,而10月地下部分养分库高于地上部分。这可能是因为养分库为生物量与养分浓度乘积,一方面,10月时地下部分有明显的生物量增加现象,因此贡献了较高的养分库;另一方面,地上部分枯死前将养分转移给地下部分,使得养分在地下部分累积。以上结果证实了冬季养分从羊草地上部分转移到地下部分保存起来,体现了多年生草本植物的生长特性。
4 结论
养分和水分有效性的提高促进了羊草地上、地下生物量和养分的积累;生长旺期生物量和养分倾向于向地上部分分配,但随着生长季结束,生物量和养分则逐渐转移至地下部分,表现了多年生植物养分保持的特性。因此,全球气候变化改变了植物生长及其对养分的吸收和分配,进而可能会对植被-土壤系统群落和结构产生影响。
[1] Bobbink R,Hornung M,Roelofs J G M.The effects of air-borne nitrogen pollutants on species diversity in natural and semi-natural European vegetation[J].Journal of Ecology,1998,86:717-738.
[2] 詹学明,李凌,浩程维信.放牧和围封条件下克氏针茅草原土壤种子库的比较[J].植物生态学报,2005,29:747-752.
[3] Niu S L,Wu M Y,Han Y,etal.Water-mediated responses of ecosystem carbon fluxes to climatic change in a temperate steppe[J].New Phytologist,2007,177:209-219.
[4] 白文明,李凌浩,宋世环.内蒙古多伦农牧交错区水资源利用现状分析[J].干旱区资源与环境,2001,15:63-67.
[5] Menge M L,Field C B.Simulated global changes alter phosphorus demand in annual grassland[J].Global Change Biology,2007,13:2582-2591.
[6] 何丹,李向林,万里强,等.施用尿素当年对退化天然草地物种地上生物量和重要值的影响[J].草业学报,2009,18(3):154-158.
[7] 邹雨坤,张静妮,杨殿林,等.不同利用方式下羊草草原土壤生态系统微生物群落结构的PLFA分析[J].草业学报,2011,20(4):27-33.
[8] 黄菊莹,袁志友,李凌浩,等.羊草绿叶氮、磷浓度和比叶面积沿氮、磷和水分梯度的变化[J].植物生态学报, 2009,33(3):442-448.
[9] Tilman D.Biodiversity:population versus ecosystem stability[J].Ecology,1996,77:350-363.
[10] Toms J D,Lespearance M L.Piecewise regression: A tool for identifying ecological thresholds[J].Ecology,2003,84:2034-2041.
[11] 牛克昌,赵志刚,罗燕江,等.施肥对高寒草甸植物群落组分种繁殖分配的影响[J].植物生态学报,2006,30(5):817-826.
[12] 吴家胜,曹福亮,应叶青,等.银杏苗期施P效应研究[J].林业科学研究,2003,16(2):171-176.
[13] 杨京平,姜宁,陈杰.施氮水平对两种水稻产量影响的动态模拟及施肥优化分析[J].应用生态学报,2003,14(10):1654-1660.
[14] 朱琴,莫邦兰,张恩让.不同施氮量对辣椒生长及落花的影响[J].贵州农业科学,2008,36(1):114-115.
[15] 迟丽华.松嫩平原羊草和虎尾草生物量及钠钾元素季节性变化的研究[J].农业系统科学与综合研究,2007,23(4):430-433.
[16] 邓斌.添加氮肥对沙地樟子松幼苗生物量分配与叶片生理特性的影响[J].生态学杂志,2006,25(11):1312-1317.
[17] 王满莲,冯玉龙.紫茎泽兰和飞机草的形态、生物量分配和光合特性对氮营养的响应[J].植物生态学报,2005,29(5):697-705.
[18] 王正文,祝廷成.水淹干扰对羊草草地地上生物量影响的初步研究[J].应用生态学报,2003,14(12):2162-2166.
[19] 田永生,郭阳耀,张培栋,等.区域净初级生产力动态及其与气象因子的关系[J].草业科学,2010,27(2):8-17.
[20] 文霞,侯向阳,穆怀彬.灌水量对京南地区紫花苜蓿生产能力的影响[J].草业科学,2010,27(4):73-77.
[21] Xu Z Z,Zhou G S.Effects of water stress and high nocturnal temperature on photosynthesis and nitrogen level of a perennial grassLeymuschinensis[J].Plant and Soil,2005,269:131-139.
[22] 张艳红,曾波,付天飞,等.长期水淹对秋华柳(SalixvariegataFranch)根部非结构性碳水化合物含量的影响[J].西南师范大学学报(自然科学版),2006,31(3):153-156.
[23] 李林锋,刘新田.干旱胁迫对桉树幼苗的生长和某些生理生态特性的影响[J].西北林学院学报,2003,19(1):14-17.
[24] 孙存华.模拟干旱诱导对藜抗旱力的影响[J].应用生态学报,1999,10(1):16-18.
[25] 孙书存,陈灵芝.辽东栎幼苗对干旱和去叶的生态反应的初步研究[J].生态学报,2000,20(5):893-897.
[26] 关军锋,马春红,李广敏.干旱胁迫下小麦根冠生物量变化及其与抗旱性的关系[J].河北农业大学学报,2004,27(1):1-5.
[27] 刘长利,王文全,崔俊茹,等.干旱胁迫对甘草光合特性与生物量分配的影响[J].中国沙漠,2006,26(1):142-145.
[28] Heckathorn S A,Deluciae E H.Retranslocation of shoot nitrogen to rhizomes and roots in prairie grasses may limit loss of N to grazing and fire during drought[J].Functional Ecology,1996,10(3):396-400.
