野豌豆属4个品种(系)的染色体核型
2012-03-13张吉宇王彦荣
管 超,张吉宇,王彦荣,聂 斌
(草地农业生态系统国家重点实验室 兰州大学草地农业科技学院,甘肃 兰州 730020)
春箭筈豌豆(Viciasativa)是豆科(Leguminosae)野豌豆属(Vicia)一年生或越年生草本植物。普遍种植于我国长江中下游、华北和西北诸省区[1]。耐干旱、耐贫瘠,不耐盐碱和水渍,喜冷凉气候,宜早春和晚秋生长,与其他作物间种或套种时有一定的耐阴性。茎枝柔嫩、叶量多,适口性好、营养价值高,是优良的绿肥、饲草和粮食兼用作物[2]。
自1998 年以来,兰州大学草地农业科技学院以早熟、高产为指标,开展了春箭筈豌豆品种选育[3],至今已成功培育出品种“兰箭3号”、品系2556和2560。这3个品种(系)的农艺学性状均具有一定的差异[4],存在基因型与环境稳定互作[5]。目前国内外学者已在狭叶野豌豆(V.angustifolia)[6-7]、广布野豌豆(V.cracca)[8]、山野豌豆(V.amoena)[9]及褐毛野豌豆(V.pannonica)[10]等多种野豌豆属植物中开展过染色体核型研究,但在春箭筈豌豆中的研究仅见于两个亚种V.sativasubsp.sativa和V.sativasubsp.nigra[11]。已有关于春箭筈豌豆种子颜色遗传[12]和细胞遗传学[13]的相关研究,但关于春箭筈豌豆不同品种(系)间染色体遗传多样性的研究鲜见报道。鉴于此,本研究对不同品种(系)的春箭筈豌豆进行染色体核型分析,深入探究不同品种(系)春箭筈豌豆的染色体核型特征和染色体遗传多样性,以期为利用染色体资料指导植物同源染色体加倍、种属间杂交、人工异源多倍体合成等技术改良或培育植物新品种提供细胞学和遗传学依据。
1 材料与方法
1.1材料及来源 供试材料包括兰州大学草地农业科技学院选育的春箭筈豌豆品种“兰箭3号”,品系2556、2560,甘肃省广为种植的狭叶野豌豆品种333/A作为对照。春箭筈豌豆品种(系)种子为兰州大学草地农业科技学院生产,333/A由中国农业科学院兰州畜牧与兽药研究所提供。
1.2方法 将成熟的野豌豆属种子放入培养皿中,加适量的水置于25 °C培养箱内生根,待幼根长约1 cm时于09:00-10:00取材;采用低温预处理的方法[14],在4 °C条件下处理24 h;预处理后的幼根于卡诺液(无水乙醇∶冰乙酸=3∶1)中固定20~24 h;固定后的幼根用去离子水反复冲洗后,放入1 mol·L-1的盐酸中,在60 °C下解离10 min;经解离的幼根用去离子水反复冲洗,再用改良的石炭酸品红染液染色0.5~2 h后压片,用普通光学显微镜(Leica,Germany)镜检;对所得图像用Photoshop 5.0进行处理分析。核型分析采用李懋学和陈瑞阳[15]的染色体分类标准,核型不对称系数(As.K,%)按Arano[16]的方法计算,核型不对称性采用Stebbins[17]的分类标准。每个品种(系)均选取16个细胞(4个根尖个体,每个根尖中选取4个细胞),计算平均值。
2 结果与分析
2.1染色体核型分析 对染色体分散良好的细胞进行观察,并进行染色体数目统计(表1)。野豌豆属4个品种(系)的染色体数目均为2n=2x=12,二倍体,染色体基数为x=6。4个不同品种(系)的染色体核型参数见表2,染色体形态、核型图和核型模式图(Idiogram)见图1。兰箭3号的染色体数为2n=12,核型公式为2n=2x=8m+2sm+2st,在6对染色体中,第1、2、4、5对染色体为中部着丝点区染色体(m),第3对染色体为近中部着丝点区染色体(sm),第6对染色体为近端部着丝点区染色体(st),未观察到次缢痕和随体;品系2556、2560的染色体数为2n=12,核型公式为2n=2x=10m+2sm,在6对染色体中,第1~5对染色体均为中部着丝点区染色体(m),第6对染色体为近中部着丝点区染色体(sm),未观察到次缢痕和随体;品种333/A的染色体数为2n=12,核型公式为2n=2x=2m+8sm (2SAT)+2st,在6对染色体中,第1、2、3、5对染色体均为近中部着丝点区染色体(sm),第4对染色体为中部着丝点区染色体(m),第6对染色体为近端部着丝点区染色体(st),在3号染色体上的长臂端观察到有1个随体存在。
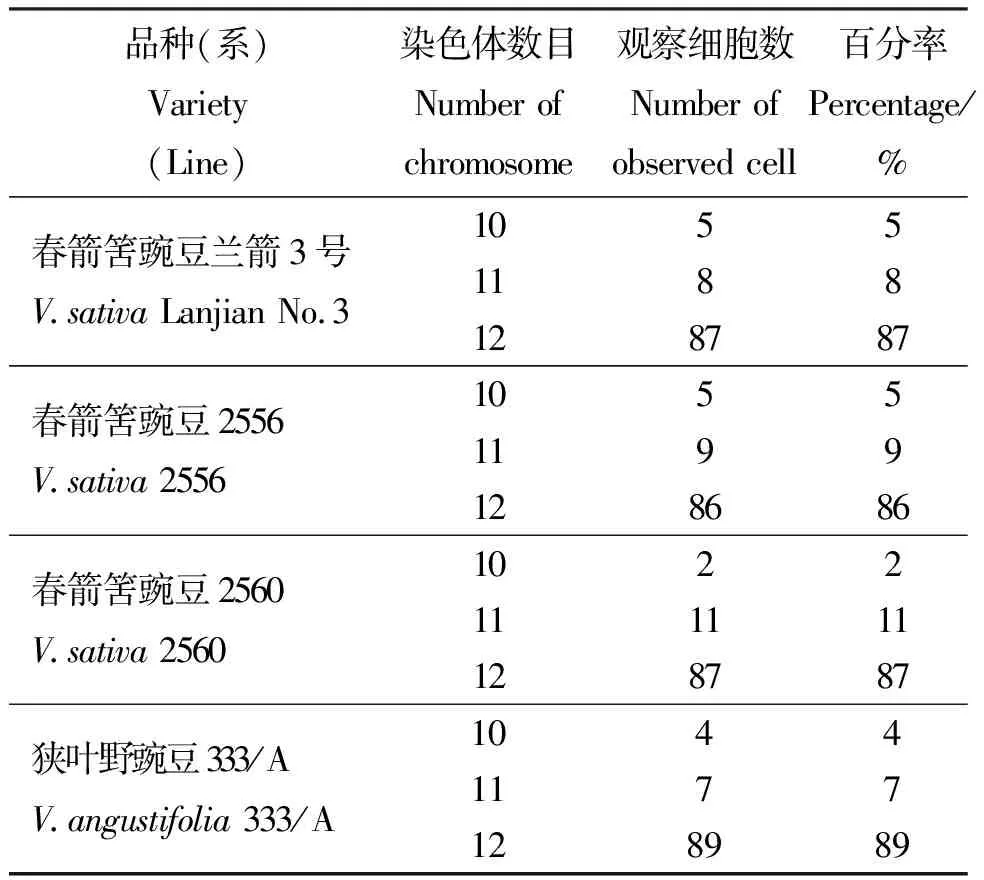
表1 野豌豆属4个品种(系)的染色体数目统计Table 1 Number of chromosomes in four Vicia varieties (lines)
2.2野豌豆属不同品种(系)的核型参数比较 根据Stebbins[17]的染色体核型分析和核型进化方法,计算染色体长度比及染色体臂比大于2∶1的染色体所占百分比,由此判断染色体核型类型。对4个品种(系)的核型参数进行比较(表3),结果显示,虽然4个品种(系)的染色体数目相同,但其核型公式、染色体总长度、长度范围、核型类型均不相同。兰箭3号和品种333/A的染色体总长度分别为41.95和50.55 μm,而品系2556和2560的染色体总长度分别为44.11和43.07 μm。兰箭3号和品种333/A的染色体绝对长度范围分别为1.84~4.78和2.44~5.68 μm,而品系2556和2560的染色体绝对长度范围分别为1.71~5.21和2.02~5.10 μm。兰箭3号的染色体的长度比为2.60,臂比>2∶1的染色体占17%,故核型类型为“2B”;品系2556的染色体的长度比为3.04,臂比>2∶1的染色体占17%,故核型类型为“2B”;品系2560的染色体长度比为2.52,不存在臂比>2∶1的染色体,故核型类型为“1B”;品种333/A的染色体的长度比为2.33,臂比>2∶`1的染色体占83%,故核型类型为“3B”。兰箭3号、品系2556、品系2560和品种333/A的核型不对称系数分别为58.64%、58.49%、58.97%和70.15%。
3 讨论
根据Stebbins[17]的染色体核型分析方法,野豌豆属4个品种(系)染色体核型有“1B”、“2B”和“3B”3种类型,表明这4个品种(系)在进化过程中处于较为进化的地位。野豌豆属4个品种(系)的染色体数目相同,均为2n=2x=12,未发生染色体数目改变,Sevimay等[18]的研究发现,8个土耳其春箭筈豌豆品种的染色体数目均为2n=2x=12,Gaffarzade等[11]研究发现,春箭筈豌豆的亚种Viciasativasubsp.sativa和V.sativasubsp.nigra的染色体数目均为2n=2x=12,而毛野豌豆(V.villosa)的染色体数目为2n=2x=14,这表明在进化过程中野豌豆属内植物发生了染色体数目的变异,而春箭筈豌豆未发生染色体数目变异。狭叶野豌豆品种333/A的核型特征与3种春箭筈豌豆的差异较大,其核型类型是相对最进化的。品系2556和2560的核型特征比较相近,二者核型公式相同,染色体总长度、长度范围和核型不对称系数都比较接近,但二者的核型类型不相同,其中品系2556较为进化。一个物种的核型特征即染色体数目、形态及行为的稳定是相对的,种内染色体的多态性是广泛存在的现象,遗传多样性越丰富,物种进化的潜力就越大;对环境改变的适应能力越强,意味着物种自身的延续能力越强[19]。本研究和前人的研究结果共同表明,不同品种(系)春箭筈豌豆的核型特征存在着广泛的差异,具有广泛的遗传多样性,表明春箭筈豌豆这个物种在进化中具有很大的潜力。
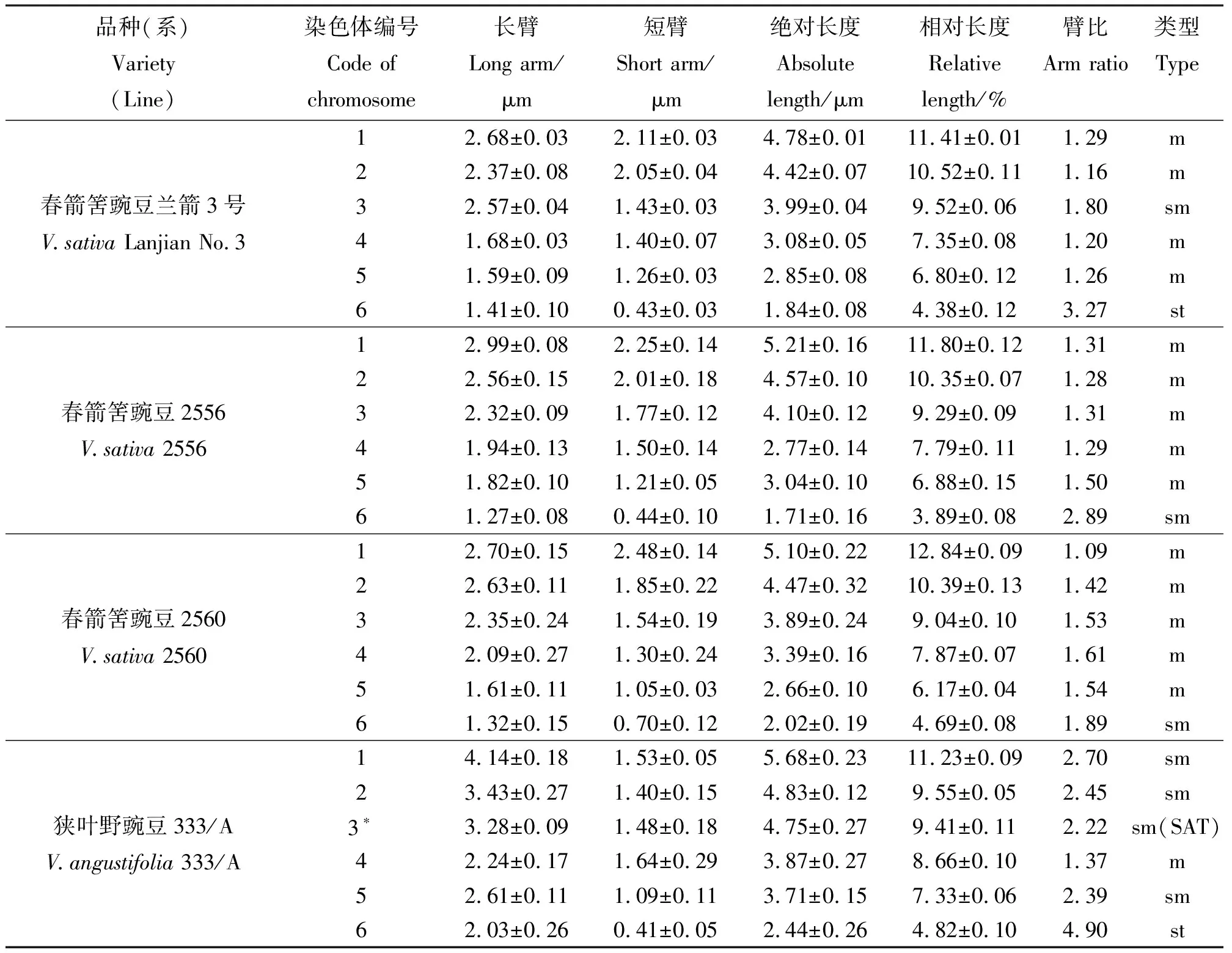
表2 野豌豆属4个品种(系)的染色体核型参数Table 2 Chromosomal parameters of four varieties (lines) of Vicia karyotype analyses
陈军和高清祥[7]发现,春箭筈豌豆品种333和333/A的核型公式均为2n=2x=2m+6st(2SAT)+4t,王建波等[6]发现狭叶野豌豆的核型公式为2n=2x=2sm+10st(2SAT),Frediani等[20]发现春箭筈豌豆的核型公式为2n=2x=2m+6sm(2SAT)+4st,而本研究发现春箭筈豌豆“兰箭3号”的核型公式为2n=2x=8m+2sm+2st,品种333/A的核型公式为2n=2x=2m+8sm(2SAT)+2st,品系2556和2560的核型公式均为2n=2x=10m+2sm。表明野豌豆属不同品种(系)在进化中发生了染色体形态上的变异,具有染色体遗传多样性。根据Arano[16]关于核型进化的观点,核型不对称系数越大,染色体越不对称,物种越进化。4个品种(系)的核型不对称系数为品种333/A>品系2560>兰箭3号>品系2556,所以说品种333/A是相对进化的。Stebbins[17]关于核型进化的分类标准为:“1A”型为最对称型,“4C”型为最不对称型,且核型比不对称者较为进化。品种333/A的核型类型为“3B”,是较为进化的类型,“兰箭3号”、品系2556的核型类型均为“2B”,品系2560的核型类型为“1B”,是较为原始的类型。陈军和高清祥[7]发现春箭筈豌豆品种333和333/A的核型类型均为“3B”,Frediani等[20]发现春箭筈豌豆的核型类型为“3B”,这均与本研究中狭叶野豌豆品种333/A的核型类型一致,而王建波等[6]发现狭叶野豌豆的核型类型为“4B”。表明野豌豆属在适应环境的过程中,产生了多种变异,经过长期的自然选择,朝着更为适应环境的方向进化。
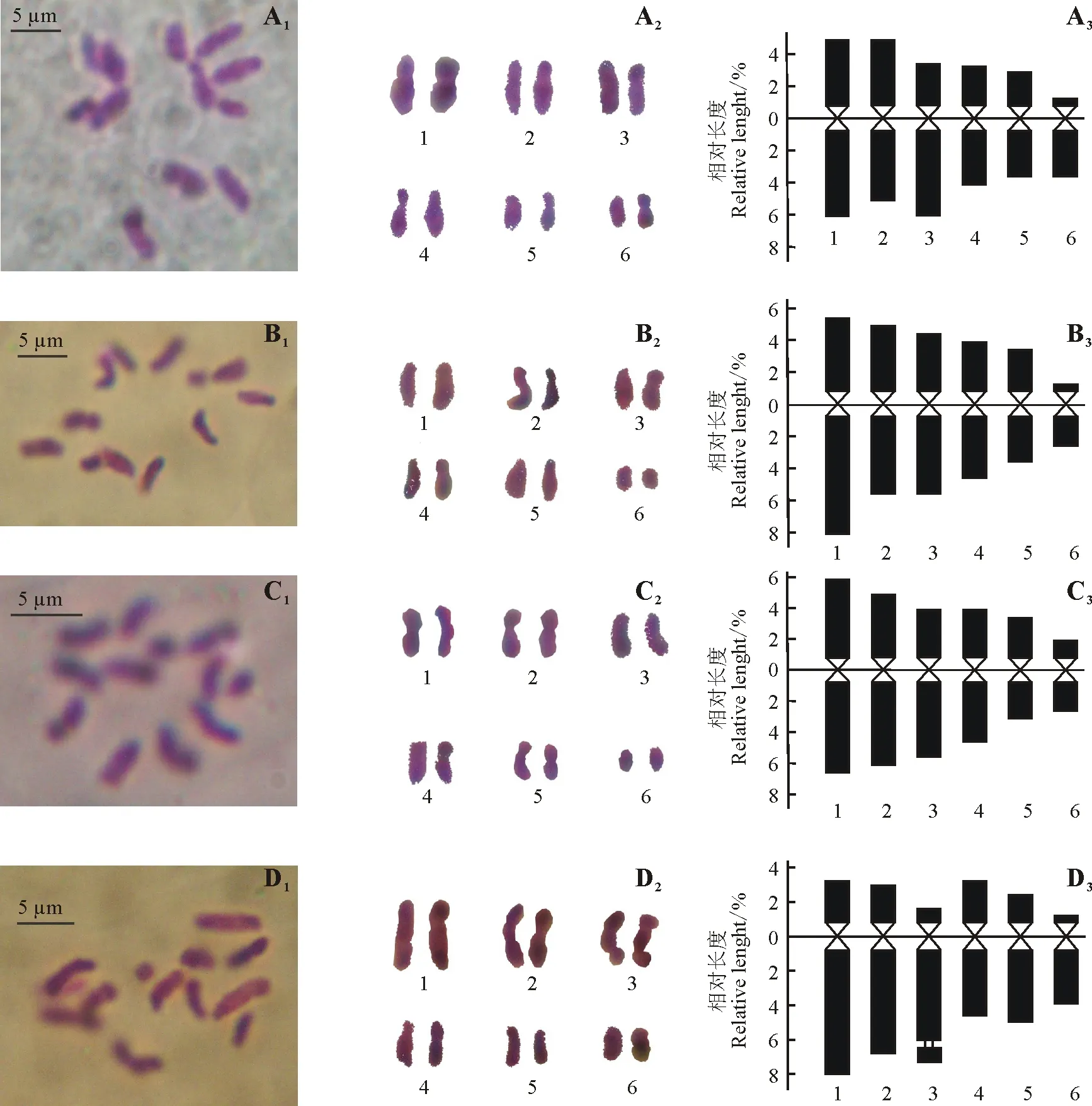
图1 兰箭3号(A1~A3)、品系2556 (B1~B3)、品系2560(C1~C3)和品种333/A(D1~D3)的染色体形态、核型图和核型模式图Fig.1 Morphology of somatic chromosomes and karyotypes, idiograms of V.sativa Lanjian No. 3 (A1-A3), V.sativa 2556 (B1-B3), V.sativa 2560(C1-C3) and V.angustifolia 333/A (D1-D3)
Alice等[10]发现春箭筈豌豆第2、3对染色体长臂端具随体,Gaffarzade等[11]发现春箭筈豌豆的亚种V.sativasubsp.sativa第2、3对染色体长臂端具随体,亚种V.sativasubsp.nigra第5对染色体长臂端具随体,陈军和高清祥[7]发现第3对染色体长臂端均具有随体,而本研究发现狭叶野豌豆品种333/A第3对染色体长臂端具随体,与前人的研究基本一致。另外,Alice等[10]还发现大花野豌豆(V.grandiflora)第2对染色体短臂端具随体,褐毛野豌豆(V.pannonica)第1、2对染色体长臂端具随体,法国豌豆(V.narbonensis)第4对染色体长臂端具随体。从理论上讲,无随体的染色体核型是不能成立的,因为随体和核仁组成区(NOR)并存,而没有NOR的细胞是不能生存的[21]。本研究在兰箭3号、品系2556和2560染色体的研究中未发现随体,是由于随体太小无法分辨,还是在制片过程中丢失,有待进一步的研究。另外,在Sevimay等[18]的研究中,8个土耳其春箭筈豌豆品种也均未发现随体的存在。
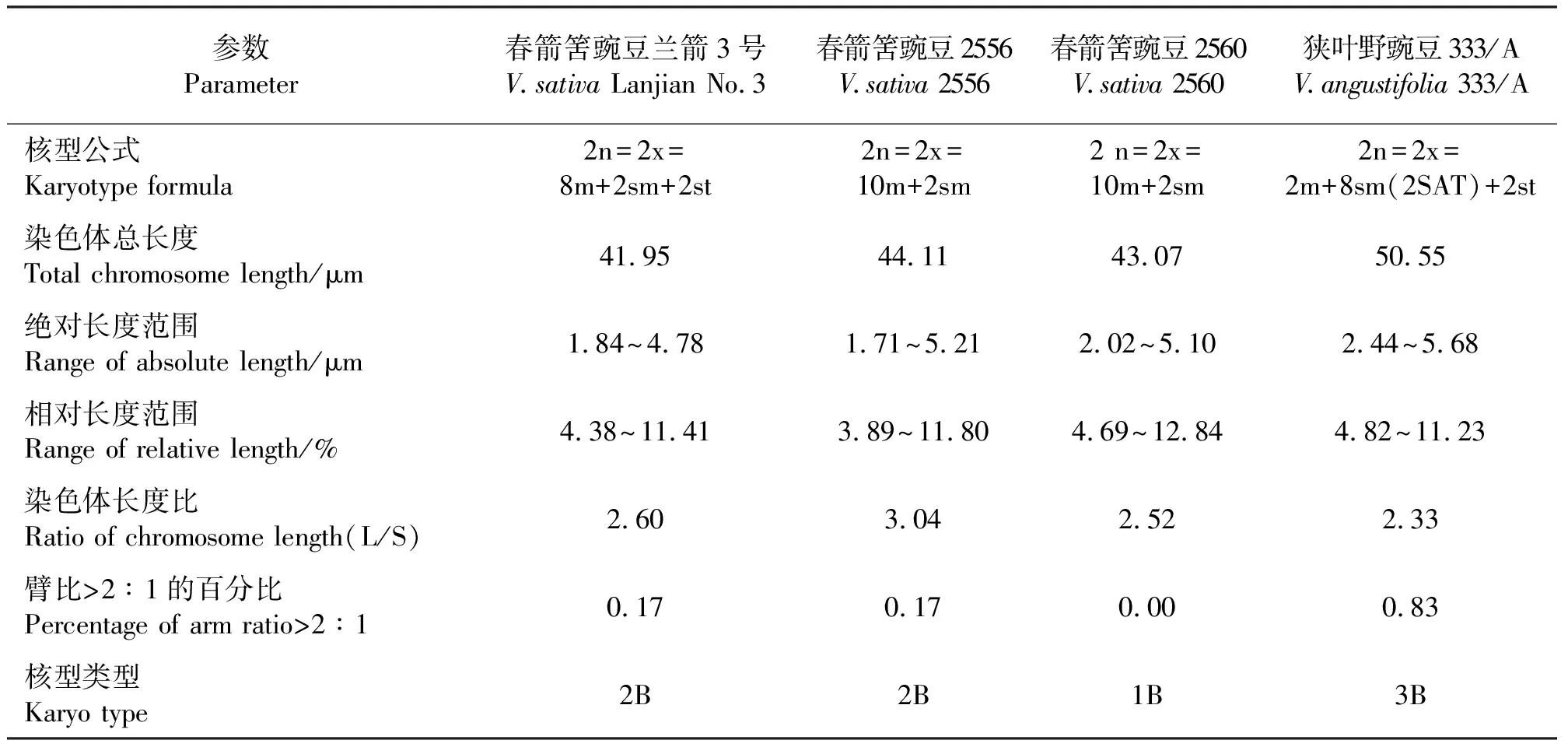
表3 野豌豆属不同品种(系)的核型参数比较Table 3 The karyotypic characters of different varieties (lines) Vicia
核型似近系数(Karyotype Resemblance-near Coefficient)是两个物种在形态结构上等同程度的表征,它反映物种间在核型上的同源性或亲缘关系的远近。谭远德和吴昌谋[22]于1993年提出了核型似近系数聚类分析。1996年吴昌谋[23]又将此分析方法进一步完善,提出了核型进化距离的概念及计算方法,2005年李峰和潘沈元[24]开发了可运行于PC机上的核型似近系数聚类分析软件,这是一种根据核型似近系数的聚类分析方法,利用Visual Basic编制的核型似近系数的聚类分析软件。因此,今后可以考虑采用核型似近系数分析软件对不同品种(系)春箭筈豌豆进行染色体核型比较,以期更准确地研究不同品种(系)春箭筈豌豆间的相关性和在进化中的关系。
遗传多样性的来源主要包括形态学性状、染色体、蛋白质、DNA等不同层次的变异,是生物多样性的基础和最重要的成分。已有关于箭筈豌豆形态学性状[4]和RFLP分子标记遗传多样性[25]的相关研究,因此,今后可以对不同品种(系)春箭筈豌豆进行DNA分子水平变异的相关研究,如DNA序列分析、随机扩增多态DNA分析(RAPD)、微卫星 DNA(Microsatellite DNA)分析、单链构象多态DNA分析(SSCP)、单核苷酸多态性(SNP)等;另外,随着测序新技术的发展,可开发春箭筈豌豆进行特异分子标记,这可进一步加速春箭筈豌豆新品种的培育。
[1] 陈默君,贾慎修.中国饲用植物[M].北京: 中国农业出版社,2002:673-675.
[2] van de Wouw M,Maxted N,Chabane K,etal.Molecular taxonomy ofViciaser.Viciabased on amplified fragment length polymorphisms [J].Plant Systematics and Evoltion,2001,229:91-105.
[3] 王赟文,南志标,王彦荣,等.高山草原条件下一年生豆科牧草生产性能的评价[J].草业学报,2001,10(2):47-55.
[4] 王彦荣,南志标,聂斌,等.几种抗寒春箭舌豌豆新品系的形态特异性比较[J].草业学报,2005,14(2):28-32.
[5] 南志标,张吉宇,王彦荣,等.五个箭筈豌豆品系基因型与环境互作效应及农艺性状稳定性[J].生态学报,2004,24(3):395-401.
[6] 王建波,曾子申,利容千.武汉地区野豌豆属3种植物的染色体研究[J].武汉植物学研究,1988,6(1):1-7.
[7] 陈军,高清祥.两个箭筈豌豆品种核型的比较研究[J].兰州大学学报(自然科学版),1993,29(2):113-116.
[8] 扈廷茂,王汉屏,张寿洲.内蒙古野豌豆属7种2变种染色体核型研究[J].内蒙古大学学报(自然科学版),1992,23(1):112-118.
[9] 苏坤梅,杨德奎.山野豌豆的染色体数目及核型分析[J].山东科学,2001,14(3):48-49,60.
[10] Alice N,Pavel N,Jiri M.Karyotype analasis ofVaciaspecies usinginsituhybridization with repetitive sequences[J].Annals of Botany,2003,91:921-926.
[11] Gaffarzadeh N L,Badrzadeh M,Asghari Z R.Karyotype of severalViciaspecies from Iran[J].Asian Journal of Plant Sciences,2008,7(4):417-420.
[12] Chowdhury D M,Rathjen J M,Tate M E,etal.Genetics of colour traits in common vetch (ViciasativaL.)[J].Euphytica,2004,136:249-255.
[13] Schifino-Wittmann M T.The cytogenetics and evolution of forage legumes from Rio Grande do Sul: A review[J].Genetics and Molecular Biology,2000,23(4):989-995.
[14] 邱希慈.植物根尖染色体的压片法[J].生物学教学,1981,1:44-45.
[15] 李懋学,陈瑞阳.关于植物核型的标准化问题[J].武汉植物学研究,1985,3(4):297-302.
[16] Arano H.Cytological studies in subfamily Carduoideae (Com-npositae) of Japan [J].IX.Botanical Magazine (Tokyo),1963,76:32-39.
[17] Stebbins G L.Chromosomal Evolution in Higher Plants[M].London: Edward Amold,1971:85-105.
[18] Sevimay C S,Guloglud D,Khawar K M.Karyotype analysis of eight Turkish vetch (ViciasativaL.) cultivals [J].Pakistan Journal of Botany,2005,37(2):313-317.
[19] 施立明.遗传多样性及其保存[J].生物科学信息,1990,2(4):158-164.
[20] Frediani M,Caputo P,Venora G,etal.Nuclear DNA contents,rDNAs,and karyotype evolution inViciasubgenusVicia: II.section Peregrinae [J].Protoplasma,2005,226:181-190.
[21] 张赞平,侯小改.杨山牡丹的核型分析[J].遗传快报,1996,18(5):63-61.
[22] 谭远德,吴昌谋.核型似近系数的聚类分析方法[J].遗传学报,1993,20(4):305.
[23] 吴昌谋.核型似近系数和进化距离的估计[J].动物分类学报,1996,21(3):338.
[24] 李峰,潘沈元.核型似近系数的聚类分析软件设计[J].徐州师范大学学报(自然科学版),2005,24(3):64-67.
[25] Potokina E,Blattner F R,Alexandrova T,etal.AFLP diversity in the common vetch (ViciasativaL.) on the world scale [J].Theoretical Applied Genetics,2002,105:58-67.
