祁连山高寒草甸生态系统昆虫群落特征
2012-03-13李婧梅陈懂懂徐世晓赵新全
蔡 海,李婧梅,程 茜,褚 晖,陈懂懂,徐世晓,赵新全,赵 亮
(1.中国科学院西北高原生物研究所,青海 西宁 810001;2.中国科学院研究生院,北京 100001;3.青海师范大学数学系,青海 西宁 810008)
生物多样性是人类赖以生存和发展的基础,草地生物多样性作为草地生态系统的重要组成部分,也是草地生态系统可持续发展的保障。昆虫是草地生物多样性的重要组成部分,它们种类众多,数量巨大,分布广泛,与脊椎动物相比,昆虫在自然界中占据了多样性更高空间尺度更小的生境,对生境的变化更敏感,具有广谱的生物地理学和生态学探针功能。近年来,随着全球气候变暖、人类活动的空前加剧和青藏高原目前正在发生的环境变迁,使得高寒草甸生物多样性发生了根本性的变化,原有植物越来越少,次生退化植物大量涌现,草地生态平衡遭到破坏[1]。生境的变化,必然会导致昆虫群落特征和生物多样性发生变化。
另外,昆虫多样性在生物多样性保护研究中占有重要地位。昆虫群落特征及多样性研究多集中在自然保护区、农田、林地、果园等区域[2-5],对草地整体区域的昆虫多样性,全面系统的研究工作尚不多见。对于中国科学院海北生态系统定位站昆虫群落的研究,早在20世纪80年代和90年代,吴亚和金翠霞[6-7],郑乐怡等[18]对昆虫群落特征和昆虫区系有过一定的研究。然而最近十几年,这方面的研究比较少。为了近一步揭示全球变化对高寒草甸昆虫多样性的影响,对祁连山地区高寒草甸的昆虫群落特征和生物多样性进行研究非常有必要。本研究通过与前人的研究结果相互比较,旨在进一步了解生境的变化对于昆虫群落特征和多样性的影响作用。
1 研究区及取样生境的自然概况
中国科学院海北高寒草甸生态系统研究站位于青藏高原东北隅的祁连山谷地,青海省海北藏族自治州门源回族自治县境内的风匣口。地处37°29′~37°45′ N,101°12′~101°23′ E。站区山地海拔4 000 m,谷地2 900~3 500 m。站区属于典型的高原大陆性气候,夏季受东南季风气候而冬季受西伯利亚寒流的影响,一年中无明显的四季之分,暖季短暂而凉爽,冷季寒冷而漫长。年平均气温-1.7 ℃,年极端最高气温27.6 ℃,极端最低气温-37.1 ℃,年降水量426~860 mm,其中80%分布于植物生长季的5-9月。全年日照时数4 434.7 h(闰年为4 445.4 h)[9]。植被类型为青藏高原典型的地带性植被-嵩草属(Kobresia)植物为建群种的高寒草甸、以西藏嵩草(K.tibetica)为建群种的沼泽化草甸和金露梅(Potentillafruticosa)为建群种的高寒灌丛草甸。高寒草甸主要分布在山地阳坡和滩地;沼泽化草甸主要分布于乱海子湖滨周围地平地段及超河漫滩低湿地段;灌丛草甸主要分布在冷湿的山地阴坡。群落结构简单、种类组成较少,植物生长期短、生物生产力较低[10]。
气候、土壤和地形是影响动物分布的重要因素外,植被也是重要的条件之一。因此,根据地形、植被和水分状况,将定位站附近地区大致分为几种不同的生境,即滩地草甸(Typical meadow)、山坡草甸(Slope meadow)、沼泽化草甸(Swamp meadow)和金露梅灌丛(Dasiphora fruticosa shrubland)。于2009年7月中旬,分别在上述4个生境进行昆虫百网取样。各生境的自然环境概况见表1。

表1 不同草地类型的自然环境概况
2 研究方法
2.1室外采样方法 采用百网取样法:使用口径为25 cm的捕虫网在样地来回100个复网(每180°扫一次为1网)捕捉昆虫[11]。
2.2室内实验方法 标本处理:大型昆虫带回室内针插,小型昆虫浸泡在75%的酒精中,均在室内计数,小型昆虫解剖镜下计数鉴定。
鉴定所需主要仪器:体式解剖镜(Motic K-400L体视显微镜)。
鉴定依据:主要依据昆虫的外部形态结构,结合检索图谱进行分类鉴定[8,12-19]。此外寄主植物、采集地也是鉴定时常依据的两个方面。
2.3数据处理方法 选取优势度指数(C)、丰富度指数(R)、多样性指数(H′)和均匀度指数(E)[20-21]。
式中,Pi为第i类群的个体数比例,即Pi=Ni/N,Ni为每一类群的个体数。N为总个体数,S为类群数。I为Mountford相似指数,a为A生境种类数,b为B生境种类数,j为A、B两生境中共同的种类数。
数据处理及绘图采用Excel 2003和Origin 8.0(群落结构取样统计与比较图、多样性计算与比较图、相似性系数)及SAS 8.0(聚类分析)。
3 结果
3.1群落类型 滩地草甸群落百网值取样获得352头昆虫,分别隶属于8目38科56种。种数最多,但个体数相对少,种类优势不明显。其中数量最多的是双翅目(Diptera)昆虫,约占36.08%,主要有麦粗腿干蝇(Meromyzasaltatrix)、背点伊蚊(Aedesdosalis)、尸蓝蝇(Cynomyamortuorum)等。膜翅目(Hymenoptera)昆虫占第2位,约占23.58%,有长凹头蚁(Formicalongiceps)、黑高加索蚁(Formicatranskallcasica)、铺道蚁(Tetramoriumcaespitum)等。居第3位的是鞘翅目(Coleoptera)昆虫,占17.33%,有蜉金龟(Aphodiussp.)、疏刻绿芫菁(Lyttabattonii)等。
山坡草甸群落百网取样获得221头昆虫,分别隶属于8目22科31种,种数和个体数均居第2位。其中双翅目种类和数量最多,约34.39%,主要有秽蝇(Coenosiasp.)、黑斑蝇(Herinasp.)等。膜翅目昆虫居第2位,约占26.24%,有长凹头蚁(Formicalongiceps)、黑高加索蚁、铺道蚁等。居第3位的是半翅目(Hemiptera),约占14.48%,主要有小长蝽(Nysiussp.)、琴长蝽(Ligyrocorissylvestr)等。
沼泽化草甸群落百网取样获得129头昆虫,分别隶属于8目15科18种。该群落昆虫以双翅目最多,约占49.61%,沼大蚊(Limoniasp.)、细腹蚜蝇(Sphaerophoriasp.)的种类最丰。鳞翅目居第2位,占13.95%,主要有条小卷蛾(Alrgyroplocesp.)、蓝灰碟(Plebejusargus)。半翅目(Hemiptera)居第3位,约占13.18%,主要有泽黑跳蝽(Saldalittoralis)、黑色小网蝽(Agrmmalaetum)。
金露梅灌丛草甸群落百网取样获得206头昆虫,分别隶属于4目19科31种。昆虫组成类群以双翅目的种类为主,其中双翅目占总数约为62.62%。这一比重高于前3种生境中双翅目的比重。这主要是该地放牧牲畜多,牦牛粪堆积也多,而且温度低湿度大,分解效率低,从而造成蝇类占优势。有黑斑蝇、尸蓝蝇(Cynomyamortuorum)、黄粪蝇(Scathophagastercorariall)等。膜翅目昆虫居第2位,约占20.39%,有长凹头蚁(Formicalongiceps)、黑高加索蚁、铺道蚁等。居第3位的是鞘翅目,占12.62%,主要是金露梅跳甲(Alticasp.)。
在以上的4个生境(表2),共取得908头昆虫,隶属于10目42科60种。调查发现,该地区双翅目昆虫是最繁盛的类群,高达43.61%;居第2位的优势目为膜翅目的蜂、蚂蚁;居第3位的是鞘翅目,种类有蜉金龟和金露梅跳甲等;其次优势目依次为,半翅目的蝽类,鳞翅目的蛾、蝶类,同翅目的蚜虫、叶蝉类。
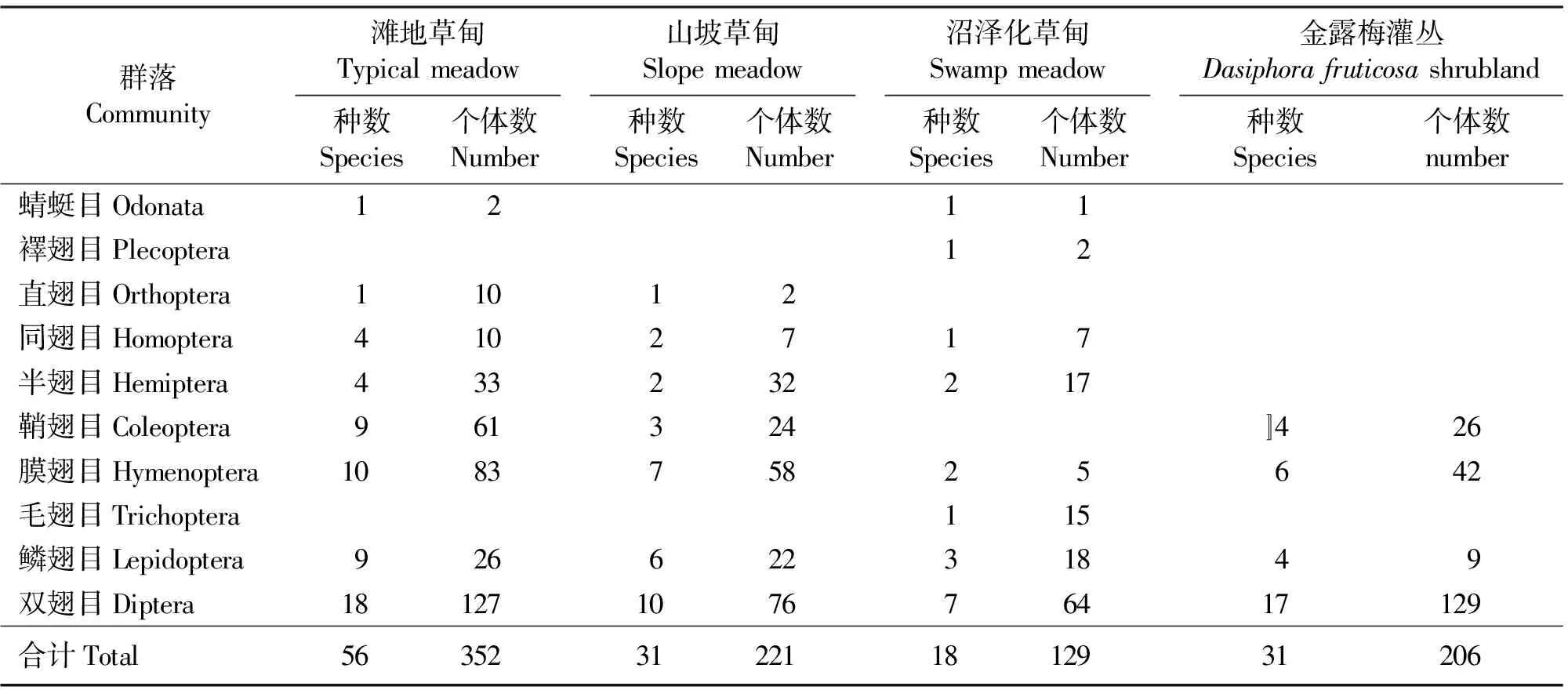
表2 高寒草甸不同生境昆虫群落取样统计(百网值)
3.2群落多样性 4种取样生境中,按多样性指数由高往低依次为滩地草甸>金露梅灌丛>山坡草甸>沼泽化草甸;群落的多样性指数和群落的丰富度指数关系密切,呈一致的趋势;优势度指数沼泽化草甸最高,滩地草甸最低;均匀性指数各群落差异较小(表3)。其中,滩地草甸不仅种类最多,而且丰富度指数也最高,因此多样性指数也最高。金露梅灌丛和山坡草甸因其丰富度和均匀性差异不大,仅次于滩地草甸,其多样性指数也较大。相反,虽然沼泽化草甸的优势度是最高的,但是因其种类较单一且数量少,其群落多样性明显低于以上3个生境群落。
3.3群落相似性与聚类分析 4个取样生境群落的相似性水平采用Mountford相似指数公式进行计算(表4),并在此基础上,采用系统聚类方法进行聚类分析,距离使用中间距离法(图1)。可以看出,4个生境中,滩地草甸和山坡草甸昆虫群落的相似性最大,这与生境相近不无关系。主要是滩地草甸和山坡草甸的植被构成较相同,只是在地形、地势上有差异。金露梅灌丛居第2位;差异最大的是滩地草甸和沼泽化草甸。

表3 不同草地类型的昆虫多样性指数
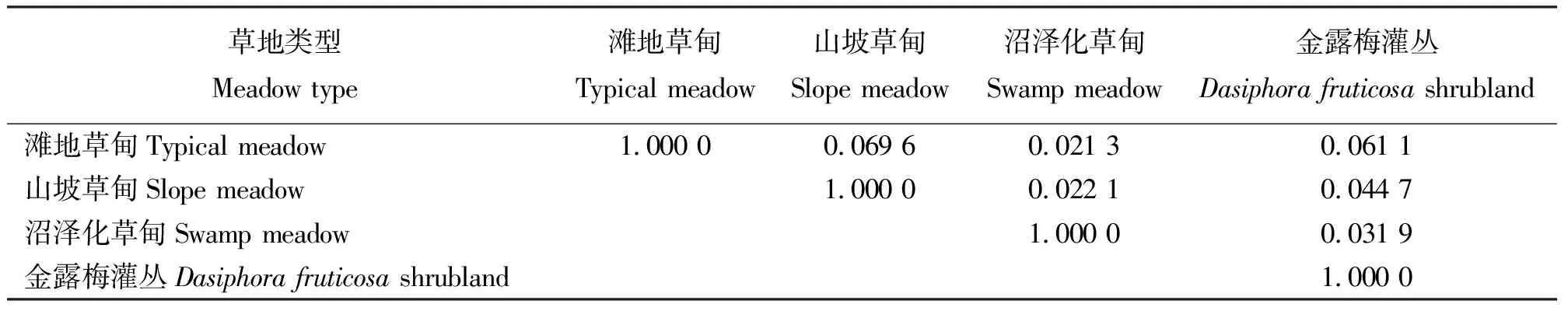
表4 不同草地类型的昆虫群落相似性

图1 高寒草甸不同生境昆虫群落相似性等级
4 讨论
4.1群落结构的变化 选取本研究各个生境的昆虫群落的优势目,与郑乐怡等[22]1990年的研究结果做对比(图2),发现各生境的优势目昆虫,都有较大的变化。滩地草甸中,2009年双翅目和半翅目昆虫个体数明显少于1990年;而膜翅目和鞘翅目昆虫的数量则多于1990年,这主要与该地区白天在畜粪堆下栖息的蜉金龟属种类较多有一定关系。山坡草甸中,2009年双翅目半翅目数量明显少于1990年;膜翅目和鞘翅目差别不大。沼泽化草甸中,2009年双翅目的数量下降的非常剧烈;其他类基本一致。金露梅灌丛中,2009年双翅目和鞘翅目数量少于1990年;鳞翅目和膜翅目差别不大。
4.2多样性的变化 选取本研究各个生境的昆虫群落多样性指数和均匀度指数,与郑乐怡等[22]1990年的结果做对比(图3),发现多样性指数与其结果基本一致,只是各生境的多样性指数均比1990年有所下降。但吴亚和金翠霞[6-7]1980和1981年所报道,沼泽化草甸的多样性最高,群落最为复杂,而据本研究观察,沼泽化草甸的植被多样性组成远低于其他3个生境,主要以西藏嵩草为优势种,昆虫种类较单一以大蚊科和摇蚊科的种类最丰,而昆虫群落的均匀度指数的变化是最为明显的,各生境基本一致而且明显高于1990年的研究结果;群落相似性与1990的研究结果有一定差异,1990年研究显著,滩地草甸与金露梅灌丛相似性最大,而本研究中滩地草甸与坡地草甸的最大,这主要与两块生境的自然环境相似有一定的关系。
群落结构和多样性指数的变化主要与全球变化和生境的变迁、退化密切相关。青藏高原是全球气候变化的敏感区和脆弱生态带,气候的微小变化均能引起高寒草甸植物群落的演替[23]。而海北站地处青藏高原的东北隅和中纬度地区,根据近20多年的气象观测资料。海北站所处的青北地区气温在逐年上升,降水却在减少。随着气温的上升,土壤的地温也在升高,这样就造成了土壤水分的散失,最终加剧干旱化程度[24-26]。土壤温湿度的变化肯定会影响到昆虫幼虫在土壤的生长发育,进而使得成虫的个体数存在差异。
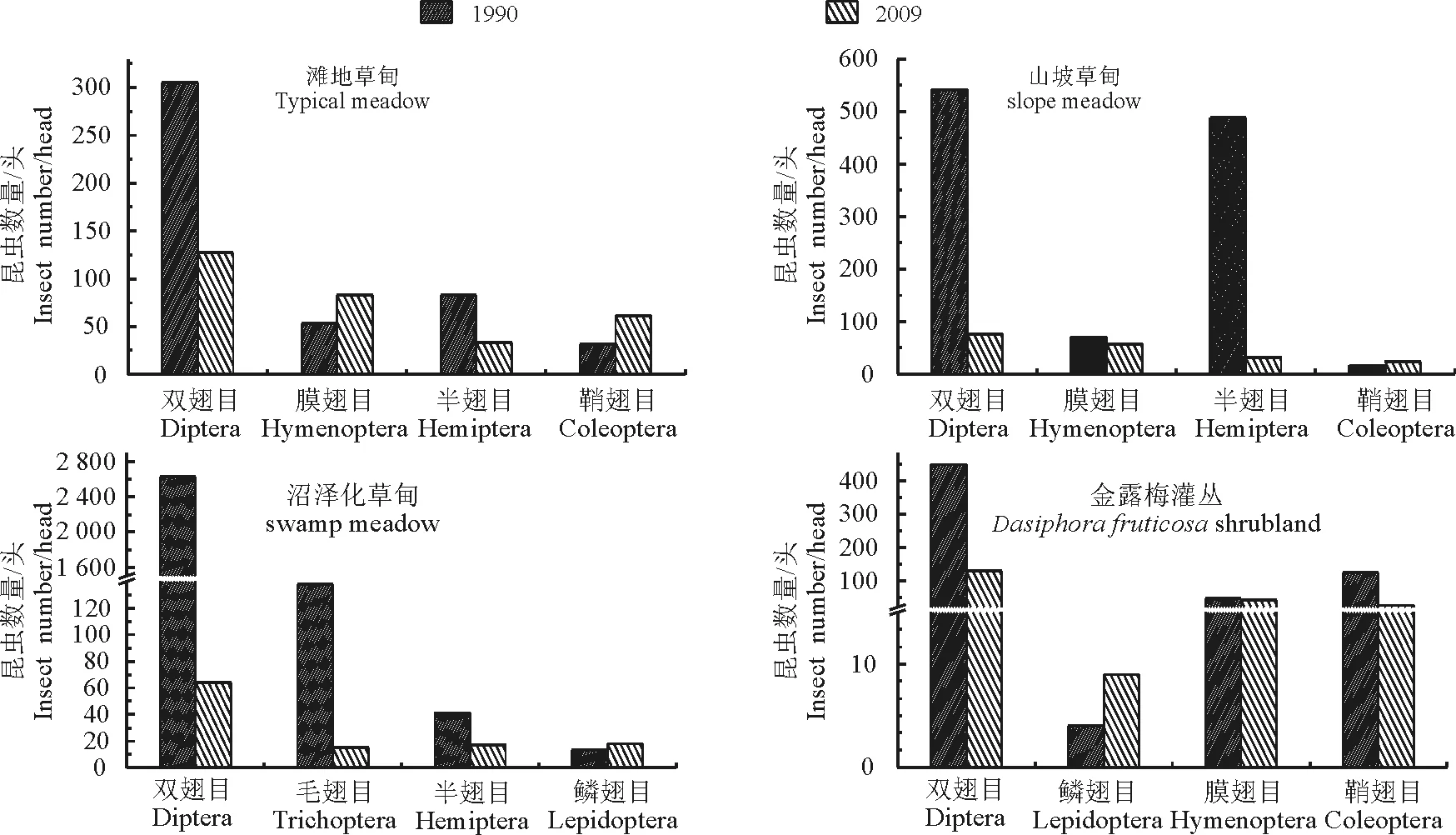
图2 不同年份各生境的昆虫结构优势目的比较
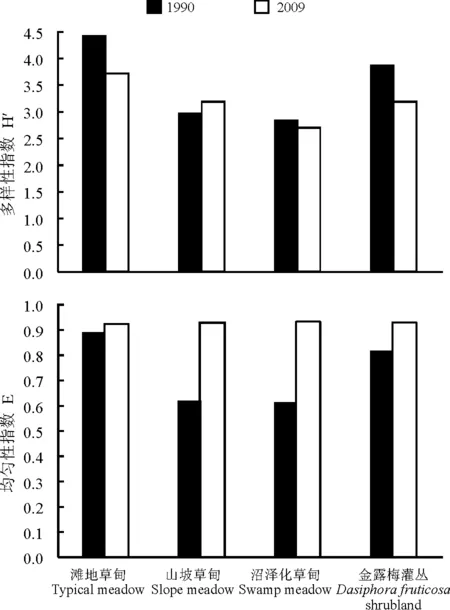
图3 不同年份各生境昆虫多样性指数的比较
近20年的过度放牧、虫鼠为害、气候变化和人为干扰,都对草地造成了一定的破坏[27-31],草地退化明显。生境面积的减少和连接度及生境间景观类型的变化,不仅影响到种群密度和种类丰度,同样也影响着植物[32-34]。通过对群落的组成、多样性和群落间的相似性分析,虽然海北定位高寒草甸地形总体上变化不大,但是在不同生境中,昆虫群落结构及其多样性存在各种程度的差异。这也进一步说明,昆虫这种小体型的动物类群对于环境变化具有较强的适应能力。生境的变迁和气候的变化引起了昆虫群落结构和多样性的差异变化。
草地生态系统是生物与环境相互作用形成的统一整体,也是人类对自然环境适应与改造形成的生态系统。昆虫作为地球上种类和数量最多的一类动物,在自然生态系统中发挥着重要作用,草地昆虫作为草地生态系统中种类和数量最多的生物组分,其功能同样不可小视,其对环境的变化极其敏感,在适应草地环境的变化过程中形成了昆虫群落结构及特征。因此,应该对不同典型草地类型的昆虫群落特征结合环境气象因子做长期的季节动态研究以及群落的演替过程,这样更能揭示昆虫对于环境变化的普遍响应机制。如何运用昆虫作为指示生物对环境的变化进行评价和监测,还有待深入研究和探讨。
[1]李希来.高寒草甸草地与其退化产物——“黑土滩”生物多样性种群群落特征的初步研究[J].草业科学,1996,13(2):21-23.
[2]孙婷婷,颜忠诚.北京地区草坪昆虫群落的初步研究[J].首都师范大学学报(自然科学版),2007,28(3):58-62.
[3]李志刚,张碧胜,龚鹏博,等.广州市昆虫群落结构及动态[J].环境昆虫学报,2009,31(3):191-196.
[4]戈峰,李典谟,谢宝瑜,等.棉田节肢动物群落的数量与能流量多样性特征分析[J].生态学报,2000,20(6):72-97.
[5]周红章,于晓东,罗天宏,等.湖北神农架自然保护区昆虫的数量变化与环境关系的初步研究[J].生物多样性,2000,8(3):262-270.
[6]吴亚,金翠霞.草甸昆虫群落及其空间与时间结构[J].昆虫学报,1980,23(2):156-166.
[7]吴亚,金翠霞.草场植被与昆虫[A].高寒草甸生态系统[C].兰州:甘肃人民出版社,1982:110-116.
[8]郑乐怡,魏美才,刘国卿.海北定位站昆虫区系初报[A].高寒草甸生态系统(第4集) [C]. 北京:科学出版社,1995:139-167.
[9]杨福屯.高寒草甸生态系统定位站自然概况[A].高原生物集刊[C].北京:科学出版社,1982:1-8.
[10]周新民,吴珍兰.植被与植物检索表——中国科学院海北高寒草甸生态系统定位站[M].西宁:青海人民出版社,2006:7-15.
[11]王新普,杨锋,杨贵军.黄河湿地银川段昆虫群落组成及其多样性研究[J].四川动物,2008,25(5):827-831.
[12]中国农业百科全书总编辑委员会.中国农业百科全书(昆虫卷) [M].北京:农业出版社,1990:208-212.
[13]牟吉元,徐洪富,荣秀兰.普通昆虫学[M].北京:中国农业出版社,1992:308-311.
[14]章士美.中国农林昆虫地理区划[M].北京:中国农业出版社,1998:208-210.
[15]中国科学院动物研究所.中国蛾类图鉴(Ⅰ、Ⅱ)[M].北京:科学出版社,1979:1-6.
[16]袁锋,张雅林,冯纪年.昆虫分类学[M].第二版.北京:中国农业出版社,2006:225-338.
[17]蔡邦华.昆虫分类学(中册)[M].北京:科学出版社,1973:56-253.
[18]朱弘复,陈一心.中国经济昆虫志(第七册:鳞翅目)[M].北京:科学出版社,1965:5-78.
[19]中国科学院动物研究所.中国农业昆虫(下册) [M].北京:农业出版社,1987:523-655.
[20]戈峰.现代生态学[M].北京:科学出版社,2005:251-257.
[21]Krebs C J.Ecological Methodology[M].Menlo Park,California:Addison Wesley Longman,Inc,1998:439-443.
[22]郑乐怡,魏美才,刘国卿.青海门源地区昆虫群落资料[A].高寒草甸生态系统(第4集)[C].北京:科学出版社,1995:127-137.
[23]赵新全.高寒草甸生态系统与全球变化[M].北京:科学出版社,2008:309-321.
[24]李英年,赵新全,曹广民,等.海北高寒草甸生态系统定位站气候、植被生产力背景分析[J].高原气象,2004,23(4):558-567.
[25]蔡英,李栋梁,汤懋苍,等.青藏高原近50年来气温的年代际变化[J].高原气象,2003,22(5):464-470.
[26]李英年.海北高寒草甸生态系统定位站近40年降水分布特征[J].资源生态环境网络研究动态,2000,11(3):9-13.
[27]李文靖,张堰铭.海北站周围3种外来物种入侵状况的初步研究[J].草业科学,2007,24(11):22-25.
[28]才旦.青海高寒草地生态系统的评价、功能失调原因和治理对策[J].草业科学,2006,23(9):7-11.
[29]崔庆虎,蒋志刚,刘季科,等.青藏高原草地退化原因述评[J].草业科学,2007,24(5):20-26.
[30]宋理明,马晓红,魏永林,等.海北州天然草地退化沙化成因与对策[J].草业科学,2009,26(7):186-190.
[31]金晓明,韩国栋.放牧对草甸草原植物群落结构及多样性的影响[J].草业科学,2010,27(4):2-10.
[32]杨芳,贺达汉.生境破碎化对生物多样性的影响[J].生态科学,2006,25(6):564-567.
[33]Fahrig L.Effects of habitat fragmentation on biodiversity[J].Annual Review of Ecology and Systematics,2003,34:487-515.
[34]Kruess A,Tscharntke T.Species richness and parasitism in a fragmented landscape:Experiments and field studies with insects onViciasepium[J].Oecologia,2000,122:129-137.
