库布齐沙地6种植物叶绿素荧光参数比较
2012-03-13闫志坚王明玖聂素梅
于 凤,高 丽,闫志坚,王明玖,聂素梅
(1.中国农业科学院草原研究所,内蒙古 呼和浩特 010010;2.内蒙古农业大学生态环境学院,内蒙古 呼和浩特 010019)
植物叶绿素荧光动力学是以叶绿素荧光作为植物体内的天然探针,快速灵敏地反映植物光合生理状况及各种外界因素对植物影响的新型、灵敏、无损伤的植物活体测定和诊断技术,被广泛应用于植物光合作用机理、环境保护、作物增产潜力预测、植物逆境生理等研究领域[1-6]。叶绿素荧光动力学技术在测定叶片光合作用过程中光系统对光能的吸收、传递、耗散、分配等方面具有独特的作用,与“表观性”的气体交换指标相比,叶绿素荧光参数更具有反映“内在性”特点[7]。近年来,在库布齐沙地普遍采用沙柳(Salixcheilophila)和优良豆科植物大面积飞播和人工栽种来防风固沙,有效改善了生态环境,增加了碳汇,所以对库布齐沙地植物生理生态适应性方面的研究也成为热点。在光合生理方面,学者们主要利用便携式光合作用系统LI-6000[8]和LI-6400[9]对生长于库布齐沙地的植物光合作用进行了研究,而利用叶绿素荧光动力学技术研究库布齐沙地植物光合作用的“内在性”还未见报道。因此,本研究以库布齐沙地东缘野生优势种油蒿(Artemisiaordosica)和大面积种植的中间锦鸡儿(Caraganaintermedia)、紫花苜蓿(Medicagosativa)、羊柴(Hedysarumlaev)、沙柳、达乌里胡枝子(Lespedezadavurica)为试验材料,通过对其叶片叶绿素荧光参数的测定,来分析6种生长于库布齐沙地的植物叶绿素荧光特性,进而探讨6种植物对沙地环境的内在适应机理,旨在为沙地植被生态恢复提供理论参考依据。
1 材料与方法
1.1研究区自然概况 研究区设在农业部鄂尔多斯沙地草原生态环境重点野外科学观测试验站中国农业科学院草原研究所十二连城试验基地,该区隶属于内蒙古鄂尔多斯市准格尔旗十二连城乡,位于库布齐沙地东缘地段,地处40°12′ N,111°05′ E。该区属于中温带大陆性季风气候,年平均气温6~7 ℃,≥10 ℃积温3 000~3 200 ℃·d,年平均降水量350~380 mm,年蒸发量2 093 mm,年日照时数为3 117 h,最高气温39.1 ℃,最低气温-32.8 ℃,无霜期145 d。植被以沙地植物为主,固定及半固定沙地以油蒿为建群种,流动沙地有白沙蒿(A.sphaerocephala)、沙蓬(Agriophyllumsquarrosum)、蓼子朴(Inulasalsoloides)等分布。土壤类型为沙壤土和风沙土。
1.2试验材料 人工种植的中间锦鸡儿、紫花苜蓿、羊柴、达乌里胡枝子、沙柳和野生优势种油蒿6种植物。
1.3研究方法 在地势平坦的固定沙地上,每种植物选择3株发育正常、长势健壮的植株,选取植株中上部且伸展方向一致的枝条,并选择枝条上第4位成熟叶片挂牌标记。所选6种植物的植株生境相同,间距均在25 m以内。2009年8月中旬选择晴朗无风天气,利用便携式叶绿素荧光分析仪(PAM-2100 Walz Germany)对标记的叶片进行活体测定。不同植物种轮回测定,测量时用黑湿布遮蔽环境光。获取荧光参数:F0(固定荧光)、Fm(最大荧光)、Fv/F0(PSⅡ的潜在活性)、Fv/Fm(PSⅡ最大光学效率或原初光能转换效率)、qP(光化淬灭系数)、NPQ(非光化学淬灭系数)、Yield(PSⅡ实际光化学效率)。
1.4试验数据统计 采用Microsoft Excel 2003软件进行试验数据的计算与图形的绘制,采用SAS 9.0统计分析软件进行方差分析和差异显著性检验。
2 结果
对植物叶绿素荧光特性变化的分析表明,6种沙地植物叶绿素荧光参数有很大差异(图1-7)。
F0值最高是油蒿,为0.297,且与其余5种植物间存在显著差异(P<0.05),而其余5种植物之间差异不显著(P>0.05),在0.278~0.283波动(图1)。
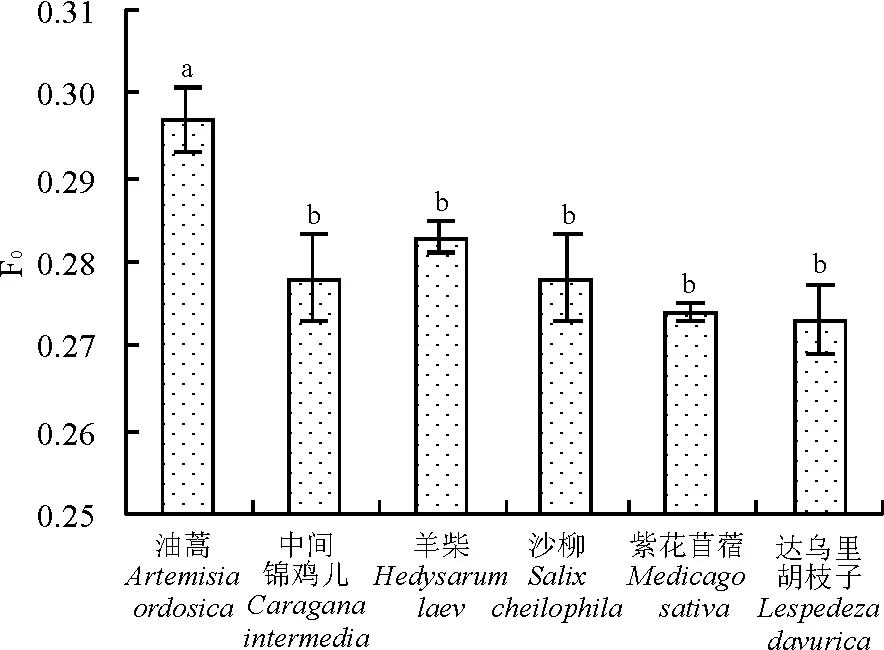
图1 6种沙地植物F0比较
Fm值排序为中间锦鸡儿>油蒿>羊柴>沙柳>紫花苜蓿>达乌里胡枝子。中间锦鸡儿具有最大Fm值,为1.751,但与油蒿、羊柴、沙柳的Fm值差异不显著(P>0.05),而显著高于紫花苜蓿和达乌里胡枝子(P<0.05),达乌里胡枝子的Fm值最小,只有1.354(图2)。

图2 6种沙地植物Fm比较
6种植物之间Fv/F0、Fv/Fm值差异性表现一致,两个参数值排序为中间锦鸡儿>沙柳>羊柴>油蒿>紫花苜蓿>达乌里胡枝子。中间锦鸡儿的Fv/F0、Fv/Fm值分别达到5.303、0.841,但与沙柳、羊柴、油蒿的Fv/F0、Fv/Fm值不存在显著性差异(P>0.05),而与紫花苜蓿和达乌里胡枝子差异显著(P<0.05)。达乌里胡枝子的Fv/F0和Fv/Fm最小,分别为3.846和0.799(图3、图4)。

图3 6种沙地植物Fv/F0比较
6种植物的qP值的排序为羊柴>中间锦鸡儿>沙柳>油蒿>达乌里胡枝子>紫花苜蓿。羊柴的qP值最高,为0.823,但与中间锦鸡儿、沙柳、油蒿的qP值差异不显著(P>0.05),而显著高于达乌里胡枝子和紫花苜蓿(P<0.05),紫花苜蓿的qP值最低,只有0.684(图5)。
6种沙地植物的NPQ值的排序为达乌里胡枝子>紫花苜蓿>油蒿>羊柴>中间锦鸡儿>沙柳。达乌里胡枝子的NPQ值最大,达到了0.471,且显著高于油蒿、羊柴、中间锦鸡儿、沙柳(P<0.05),而与紫花苜蓿差异不显著(P>0.05)(图6)。
6种植物中,Yield值的排序为羊柴>中间锦鸡儿>沙柳>油蒿>达乌里胡枝子>紫花苜蓿。羊柴的Yield值最高,为0.619,且与油蒿、达乌里胡枝子和紫花苜蓿差异显著(P<0.05),但与中间锦鸡儿和沙柳的Yield值差异不显著(P>0.05) (图7) 。

图4 6种沙地植物Fv/Fm比较

图5 6种沙地植物qP比较
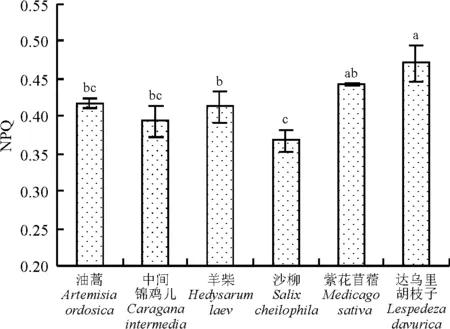
图6 6种沙地植物NPQ比较
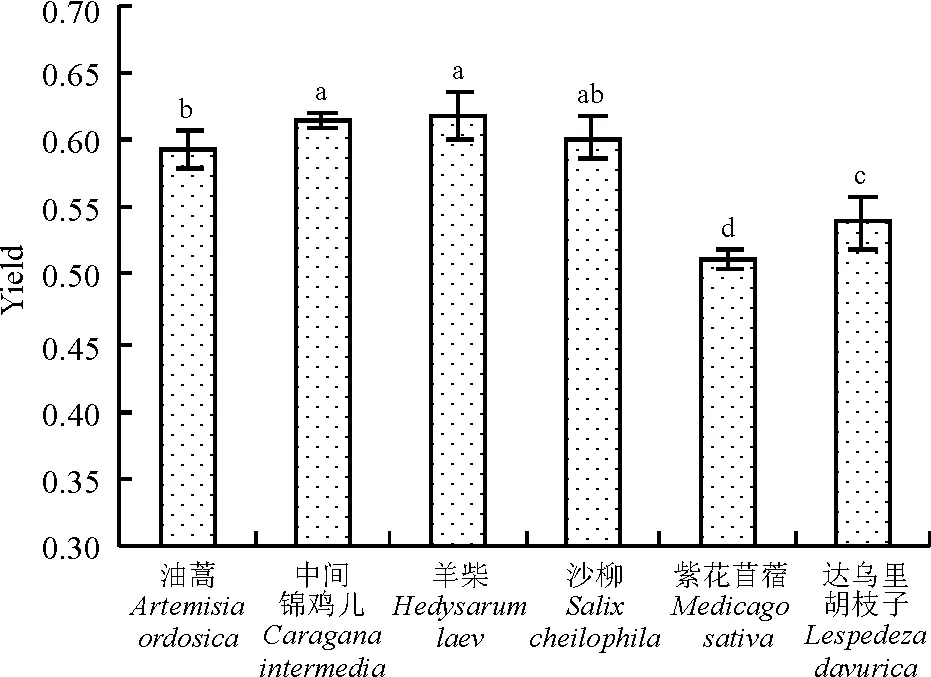
图7 6种沙地植物Yield比较
3 讨论与结论
据研究,植物的叶绿素荧光特性因种类和生态环境的不同有很大的差异,如不同阔叶树种叶绿素荧光参数差异显著[10];在遮阴与全光照下,濒危药用植物川贝母(Fritillariacirrhosa)叶绿素荧光特性有明显不同[11]。本研究得出,生长于库布齐沙地的中间锦鸡儿、紫花苜蓿、羊柴、油蒿、沙柳、达乌里胡枝子之间的F0、Fm、Fv/F0、Fv/Fm、qP、NPQ、Yield 7项叶绿素荧光参数表现出不同程度的差异。
正常情况下,叶绿素吸收的光能主要通过光合电子传递、叶绿素荧光和热耗散3种途径消化掉,这3种途径间存在着此消彼长的关系,光合作用和热耗散的变化会引起荧光发射的相应变化,因此,荧光变化可以反映光合作用和热耗散的情况[12]。Fm表示PSⅡ反应中心处于完全关闭时的荧光产量,可以反映通过PSⅡ的电子传递情况[7],Fm= F0+Fv[13],F0是固定荧光或初始荧光,也称基础荧光和0水平荧光,是光系统II(PSⅡ)反应中心处于完全开放时的荧光产量,它与叶片叶绿素浓度有关[7]。Fv代表可参与PSⅡ光化学反应的光能辐射部分,反映PSⅡ且原初电子受体QA的还原情况[13]。Fv/F0表示PSⅡ的潜在活性[7]。Fv/Fm是PSⅡ原初光能转化效率,反映植物的潜在最大光合能力[14]。本研究发现,在6种沙地植物中,中间锦鸡儿、羊柴、沙柳、油蒿4种灌木植物的Fm、Fv/F0、Fv/Fm值均显著高于草本状半灌木达乌里胡枝子和草本植物紫花苜蓿(P<0.05),说明中间锦鸡儿、羊柴、沙柳、油蒿的PSⅡ潜在活性和潜在最大光合能力均大于达乌里胡枝子和紫花苜蓿。光化学淬灭系数qP是由光合作用引起的荧光淬灭,反映植物光合活性的高低[13]。本研究结果表明,羊柴、中间锦鸡儿、沙柳、油蒿的qP值较高,且显著高于紫花苜蓿和达乌里胡枝子(P<0.05),说明羊柴、中间锦鸡儿、沙柳、油蒿的光合活性高于紫花苜蓿和达乌里胡枝子。Yeild反映PSⅡ反应中心在部分关闭情况下的实际原初光能捕获效率,能准确地反映实际的PSⅡ中心进行光化学反应的效率情况[12],体现植物的实际光合能力(实际光合效率)[14]。在所测的6种植物中,Yield值由大到小排序,羊柴>中间锦鸡儿>沙柳>油蒿>达乌里胡枝子>紫花苜蓿,说明羊柴、中间锦鸡儿、沙柳、油蒿的实际光合能力也大于达乌里胡枝子和紫花苜蓿。非光化学淬灭NPQ反映的是PSⅡ天线色素吸收的光能不能用于光合电子传递而以热的形式耗散掉的光能部分,非光化学淬灭是一种自我保护机制,对光合机构起一定的保护作用[7]。供试植物中,达乌里胡枝子和紫花苜蓿的NPQ值较高,分别为0.442和为0.471,显著高于中间锦鸡儿、羊柴、油蒿(P<0.05),说明紫花苜蓿和达乌里胡枝子的叶片通过热耗散消耗掉过剩光能,从而避免了沙地强光环境对光合机构的破坏,具有较强的光保护能力,这也可能是这两种植物能够适应沙地恶劣环境的重要原因之一。
中间锦鸡儿、羊柴、沙柳、油蒿、紫花苜蓿和达乌里胡枝子的叶绿素荧光特性表现出它们对沙地环境较强的适应性。中间锦鸡儿、羊柴、沙柳、油蒿的光合能力较强,在沙地干旱、强光环境下,它们通过光合作用充分利用光能,从而积累大量的有机物质来适应环境。紫花苜蓿和达乌里胡枝子虽然光合能力较弱,但它们能将过剩的光能用于热耗散,从而避免叶片受到伤害,达到自我保护的目的。
[1]林世青,许春辉,张其德,等.叶绿素荧光动力学在植物抗性生理生态学和农业现代化中的应用[J].植物学通报,1992,9(1):1-6.
[2]徐德聪,吕芳德,潘晓杰.叶绿素荧光分析技术在果树研究中的应用[J].经济林研究,2003,21(3):88-91.
[3]郭春芳,孙云.叶绿素荧光动力学在植物抗性生理研究中的应用[J].福建教育学院学报,2006(7):120-123.
[4]陈小凤,李杨瑞,叶燕萍,等.利用叶绿素荧光参数和净光合速率评价引进禾本科牧草的抗旱性[J].草业科学,2007,24(5):53-57.
[5]郭玉朋,郑霞,王新宇,等.叶绿素荧光技术在筛选光合突变体中的应用[J].草业学报,2009,18(6):226-234.
[6]姜英,彭彦,李志辉,等.植物生长延缓剂对金钱树抗寒性指标的影响[J].草业科学,2010,27(9):51-56.
[7]张守仁.叶绿素荧光动力学参数的意义及讨论[J].植物学通报,1999,16(4):444-448.
[8]高清竹,杨劼,乌力吉,等.库布齐沙地油蒿光合作用特性与环境因子的关系[J].中国沙漠,1999,19(3):276-279.
[9]马阔东,高丽,闫志坚,等.库布齐沙地三种植物光合、蒸腾特性和水分利用效率研究[J].中国草地学报,2010,32(2):116-120.
[10]郑淑霞,上官周平.8种阔叶树种叶片气体交换特征和叶绿素荧光特性比较[J].生态学报,2006,26(4):1080-1087.
[11]李西文,陈士林.遮阴下高原濒危药用植物川贝母光合作用和叶绿素荧光特征[J].生态学报,2008,28(7):3438-3446.
[12]Rohacek K.Chlorophyll fluorescence parameters:the definitions,photosynthetic meaning and mutual relationships[J].Photosynthetica,2002,40(1):13-29.
[13]陈建明,俞晓平,程家安.叶绿素荧光动力学及其在植物抗逆生理研究中的应用[J].浙江农业学报,2006,18(1):51-55.
[14]Schreiber U,Bilger W,Neubauer C.Chlorophyll fluorescence as a non-intrusive indicator for rapid assessment of in vivo photosynthesis[A].In:Schulze E D,Caldwell M M.Ecophysiology of Photosynthesis[M].Berlin:Springer-Verlag,1994:49-70.
