物种多样性对豆科牧草功能特征的影响
2012-03-13张灵菲葛庆征张卫国江小雷
张灵菲,魏 斌,郝 敏,葛庆征,傅 华,张卫国,江小雷
(兰州大学草地农业科技学院,甘肃 兰州 730020)
豆科牧草因其特殊的生物固氮作用,具有改善土壤氮肥供应,促进其他牧草,特别是禾本科牧草生长发育,以及较高的营养价值等特性而在天然草地和人工草地生态系统中占有重要地位[1-3]。在草地生态系统生物多样性研究中,豆科牧草作为一个特殊的功能类群,对生物多样性促进生态系统功能发挥着重要的作用[4-5]。研究表明,一方面,随着植物群落物种多样性的增加,豆科植物对生态系统功能的促进作用有所增强[3,6-8];另一方面,群落中物种多样性的增加势必会导致种间竞争作用的增强,进而不可避免地会对豆科牧草的生长发育产生负面影响。然而,物种多样性对豆科牧草的影响则少见报道[7,9-10]。
研究表明[11-12],植物的组成或种类对生态系统过程的影响本质上是通过物种的功能特征而体现的,植物的功能特征可直接影响能量流通和物质循环,间接改变非生物条件,进而调节生态系统过程[11]。关于物种多样性、干扰及环境变化对植物功能特征影响的研究有很多报道[13-15],但豆科牧草功能特征响应环境条件变化的研究尚不多见[9,16]。本研究的目的是探讨物种多样性会如何影响豆科牧草的功能特征及生产力,以期为豆科牧草的利用、人工草地的建植和管理提供有益参考。
1 材料与方法
1.1试验材料 试验用瓦盆直径为30 cm,高为30 cm。草种为8种一年生植物。其中豆科植物有箭筈豌豆(Viciasativa,Vs)、山黧豆(Lathyrussativus,Ls),禾本科植物有C3植物燕麦(Avenasativa,As)、多花黑麦草(Loliummultiflorum,Lm)和C4植物苏丹草(Sorghumsudanense,Ss)、稗(Echinochloacrusgalli,Ec),杂类草有籽粒苋(Amaranthushypochondriacus,Ah)、饲用油菜(Brassicacapestris,Bc)。
试验于2010年6月20日-8月20日在兰州大学草地农业科技学院院内采用室外盆栽法进行。播种密度:单播每种每盆均播种30粒;混播每盆的总播量不变,但各物种播量按物种数的增加而递减。每盆装过筛农田熟土8 kg。于2010年6月20日将预先发芽的种子按设计密度点播于盆内。试验设Ⅰ(8个种的单播),Ⅱ(VsAs,LsAs,VsEc,LsEc,VsAh),Ⅳ(VsAsLmSs,LsAsLmSs,VsEcAhLm,LsEcAhLm),Ⅵ(VsAsLmSsEcAh,LsAsLmSsEcAh),Ⅷ(VsLsAsL-mSsEcAhBc)5个多样性组合,共20个处理,每处理重复3次,共60 盆,采用随机区组设计。试验期间保证盆内有充足的水分(土壤含水量在30%左右),以利于牧草正常生长。所有处理均于2010年8月20日一次性收获,同时测定不同物种的功能特征及地上生物量。所有样品均于65 ℃ 条件下烘48 h测干质量。
测定的功能特征主要包括相对生长率(Relative growth rate,RGR,g · g-1·d-1) 、比叶面积(叶面积/叶干质量,cm2·g-1)、株高(cm)、根深(cm)、叶干物质含量(叶烘干质量/叶鲜质量,mg·g-1)和单位叶面积根量(根鲜质量/叶面积,g·m-2)6个与生产力、光合能力、养分利用等功能密切相关的特征。比叶面积、叶干物质含量按 Garnier 等[17]的方法测定。株高、根深和单位叶面积根量按Cornelissen等[18]的方法测定。相对生长率按Hunt[19]的方法计算:
RGR=[lg(Wf/Wi)]/Δt。
式中,Wf为植物收获时的干质量,Wi为植物的初始干质量(用各物种的种子质量表示),Δt为植物的生长天数。
1.2数据分析 采用SPSS 13.0统计分析软件对所有数据进行统计分析。使用单因素方差分析(ANOVA)对各变量在不同物种间的差异显著性进行统计检验,用简单线性相关方法对豆科植物各功能特征与物种多样性的关系进行分析。
2 结果
2.1物种多样性对豆科牧草功能特征的影响 简单相关分析表明,物种多样性对豆科牧草各功能特征有不同的影响(图1)。物种多样性对豆科牧草比叶面积、株高有促进作用,二者呈显著正相关关系(图1B、图1C),说明豆科牧草的比叶面积和株高随着物种多样性的增加而增加。而物种多样性对豆科牧草的其他功能特征则有负面影响:相对生长率(图1A)、根深(图1D)、叶干物质含量(图1E)和单位叶面积根量(图1F)4个功能特征均随物种多样性的增加而下降。

图1 物种多样性对豆科牧草功能特征的影响
2.2物种多样性对豆科牧草生产力的影响 无论豆科牧草的地上生产力(图2A)还是地下生产力(图2B)都随着物种多样性的增加而下降,表明随着物种多样性的增加,其他牧草对养分的争夺性利用影响了豆科牧草生物量的积累。
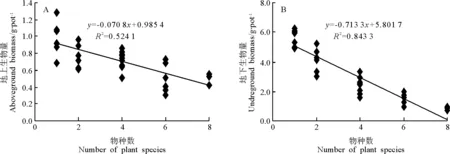
图2 物种多样性对豆科牧草生产力的影响
3 讨论
随着物种多样性的增加,不同物种间的互补作用可使植物群落对地上及地下资源的利用能力增强[6-7],进而增强生态系统功能,而豆科牧草的出现对增强多样性群落中物种间的互补作用具有关键性作用[8]。豆科牧草功能特征的变化可在一定程度上反映多样性群落中物种间的相互关系,以及对生态系统功能,特别是系统生产力的影响[20]。
本研究表明,物种多样性对豆科牧草的功能特征有重要影响,但对体现生长发育、养分利用和竞争对策等不同的功能特征,其影响模式不同。物种多样性对豆科牧草的比叶面积和株高有促进作用。比叶面积代表植物单位干质量叶的光截获面积,与植物的同化率密切相关,是重要的功能特征之一[21]。植株的高度是体现植物空间(地上)资源竞争能力的标志[22]。此结果说明,随着物种多样性的增加,群落中各物种对地上光能资源的竞争作用增强,从而刺激豆科牧草通过表型可塑性的变化,改变生存对策,增强获取地上资源的能力,进而适应多样性群落中较强的竞争作用[22]。
相对生长率是物种响应环境变化的关键性特征,相对生长率较高的物种具有相对较高的生产力水平[23]。根深是体现植物地下资源竞争能力的功能特征,对生产力有促进作用[15]。叶干物质含量代表叶组织密度,与植物养分保持能力密切相关[21]。叶干物质含量较高,植物生长较慢,具有较低的比叶面积[13]。单位叶面积根量体现植物水分吸收和水分蒸腾的平衡关系[24],较高的单位叶面积根量值意味着植物将更多的生物量分配于根的生长而较少的分配给叶,进而降低植物的相对生长率,最终导致较低的生产力。本研究中,豆科牧草的相对生长率、根深、叶干物质含量和单位叶面积根量4个功能特征值随着物种多样性的增加而减少的事实表明,在多样性较高的群落中,其他物种对资源的竞争作用在一定程度上限制了豆科牧草的生长,此结果间接地证明了Carlsson等[9]的研究,表明在多样性较高的群落中,豆科牧草因其固氮作用而对其他植物,特别是禾本科植物的生长有促进作用,但其他植物则对豆科牧草的生长及固氮作用有一定的不利影响,从而导致群落中豆科牧草地上、地下生产力水平的降低。
综上所述,物种多样性对豆科牧草各功能特征有不同的影响。物种多样性可促进豆科牧草与地上资源(主要是光能)利用相关的功能(增加比叶面积和株高),而减弱与地下资源利用相关的功能(如根深和单位叶面积根量),进而降低其生长率(减少相对生长率和叶干物质含量),最终导致豆科牧草生产力的减少。由此可见,在多样性群落中,豆科牧草一方面对促进群落资源的互补性利用有重要贡献,另一方面,其自身功能的发挥受到其他物种对有效资源的竞争性利用的限制。因此,在建立混播草地和维持草地物种多样性时,应适当考虑豆科牧草的利益,使其更好地发挥固氮功能。
[1]江小雷,岳静,张卫国,等.生物多样性,生态系统功能与时空尺度[J].草业学报,2010,19(1):219-225.
[2]秦文静,梁宗锁.四种豆科牧草萌发期对干旱胁迫的响应及抗旱性评价[J].草业学报,2010,19(4):61-70.
[3]Mulder C P H,Jumpponen A,Högberg P,etal.How plant diversity and legumes affect nitrogen dynamics in experimental grassland communities [J].Oecologia,2002,133:412-421.
[4]Spehn E M,Scherer-Lorenzen M,Schmid B,etal.The role of legumes as a component of biodiversity in a cross-European study of grassland biomass nitrogen [J].Oikos,2002,98:205-218.
[5]Dybzinski R,Fargione J E,Zak D R,etal.Soil fertility increases with plant species diversity in a long-term biodiversity experiment[J].Oecologia,2008,158:85-93.
[6]穆锋海,王梅,陈远其,等.牧草营养性状与生物量关系的研究[J].草业科学,2009,26(10):122-126.
[7]陈玮玮,万里强,何峰,等.温度和光照时间对3个秋眠型紫花苜蓿品种形态特征的影响[J].草业科学,2010,27(12):113-119.
[8]Temperton V M,Mwangi P N,Scherer-Lorenzen M,etal.Positive interactions between nitrogen-fixing legumes and four different neighbouring species in a biodiversity experiment[J].Oecologia,2007,151:190-205.
[9]Carlsson G,Palmborg C,Jumpponen A,etal.N2fixation in three perennialTrifoliumspecies in experimental grasslands of varied plant species richness and composition[J].Plant Ecology,2009,87-104.
[10]Jumpponen A,Mulder C P H,Huss-Danell K,etal.Winners and losers in herbaceous plant communities:insights from foliar carbon isotope composition in monocultures and mixtures[J].Journal of Ecology,2005,93:1136-1147.
[11]Díaz S,Hodgson J G,Thompson K,etal.The plant traits that drive ecosystems:Evidence from three continents[J].Journal of Vegetation Science,2004,15:295-304.
[12]Lavorel S,Garnier E.Predicting changes in community composition and ecosystem functioning from plant traits:revisiting the Holy Grail[J].Functional Ecology,2002,16:545-556.
[13]Garner E,Cortez J,Billés G,etal.Plant functional markers capture ecosystem properties during secondary succession[J].Ecology,2004,85:2630-2637.
[14]de Bello F,Lepš J,Sebastia M-T.Predictive value of plant traits to grazing along climatic gradient in the Mediterranean[J].Journal of Applied Ecology,2005,42:824-833.
[15]Klimešová J,Latzel V,de Bello F,etal.Plant functional traits in studies of vegetation changes in response to grazing and mowing:towards a use of more specific traits[J].Preslia,2008,80:245-253.
[16]Roscher C,Schmid B,Buchmann N,etal.Legume species differ in the responses of their functional traits to plant diversity[J].Oecologia,2011,165(2):437-452.
[17]Garnier E,Shipley B,Roumet C,etal.A standardized protocol for the determination of specific leaf area and leaf dry matter content[J].Functional Ecology,2001,15(5):688-695.
[18]Cornelissen J H C,Lavorel S,Garnier E,etal.A handbook of protocols for standardized and easy measurement of plant functional traits worldwide[J].Australian Journal of Botany,2003,51(4):335-380.
[19]Hunt R.Plant Growth Analysis [M].London:Academic Press,1997:101.
[20]Craine J M,Tilman D,Weding D,etal.Functional traits,productivity and effects on nitrogen cycling of 33 grassland species[J].Functional Ecology,2002,16,563-574.
[21]Pontes L D,Soussana J F,Louault F,etal.Leaf traits affect the above-ground productivity and quality of pasture grasses[J].Functional Ecology,2007,21:844-853.
[22]Westoby M,Falster D S,Moles,etal.Plant ecological strategies:Some leading dimensions of variation between species[J].Annual Review of Ecology,Evolution,and Systematics,2002,33(1):125-159.
[23]Vile D,Shipley B,Garnier E.Ecosystem productivity can be predicted from potential relative growth rate and species abundance[J].Ecology Letters,2006,9:1061-1067.
[24]Kudoh T T H,Kachi N.Responses of root length/leaf area ratio and specific root length of an understory herb,Pteridophyllumracemosum,to increases in irradiance[J].Plant and Soil,2003,255(1):227-237.
