亚洲小车蝗飞行能力及其与种群密度的关系
2012-03-13高书晶魏云山特木儿刘爱萍徐林波
高书晶,魏云山,特木儿,刘爱萍,徐林波,王 宁
(1.中国农业科学院草原研究所,内蒙古 呼和浩特 010010; 2.赤峰市农牧科学研究院,内蒙古 赤峰 024031;3.乌兰察布盟四子王旗草原站,内蒙古 乌兰花 011800)
亚洲小车蝗(Oedaleusasiaticus)是我国北方草原的主要优势种害虫之一,也是农牧交错地带的重要经济害虫,主要分布于内蒙古、河北、甘肃和青海等省(区)。亚洲小车蝗主要以禾本科植物为食,近年来持续爆发成灾,一般占据整个蝗虫种群数量的50%~60%,严重发生时能超过90%[1]。亚洲小车蝗的发生为害早,发生数量大,可致受害作物减产10%~30%,严重时超过50%,甚至毁种或绝收[2-3],对农牧业生产造成极大损失,因此,有效控制其发生和危害至关重要。
亚洲小车蝗飞行能力很强,近年来,亚洲小车蝗在大发生时常常出现聚集取食并且集体转移的习性,曾有人观察到其有迁飞现象,它不仅危害草原,而且群集式地在城市中心大量出现,给人们的生活也造成了极大的困扰[4]。在非洲曾出现过塞内加尔小车蝗(O.senegalensis)远距离聚集迁飞,一晚上可以迁飞350 km[5-6],亚洲小车蝗与塞内加尔小车蝗同属于斑翅蝗科小车蝗属,因此其习性有可能相近。多种研究表明,亚洲小车蝗可能具有远距离迁飞的习性,多在夜间飞行且具有较强的趋光性[7-8]。目前,国内对亚洲小车蝗的研究主要集中在发生规律、生态学以及防治方法等方面[9-10],有关其成虫飞行能力的研究甚少,而且亚洲小车蝗是否有群居型与散居型的分化,种群密度对其飞行的影响,这些都未被研究过,明确这些问题的答案可为确定亚洲小车蝗的迁飞习性及进一步研究其迁移扩散规律奠定必要的基础,并为我国亚洲小车蝗的监测预警和综合防治提供重要的理论依据,对完善和促进昆虫飞行生物学研究及迁飞学科发展等具有重要的理论和现实意义。
1 材料与方法
1.1饲养方法 亚洲小车蝗试验种群于2011年采自内蒙古四子王旗和阿拉善左旗,主要采集1-2龄蝗蝻,高密度种群主要选择亚洲小车蝗大爆发和聚集危害地区,一般500头·m-2以上,低密度种群主要选择亚洲小车蝗危害在防治指标以下的零星发生区,一般3~5 头·m-2,经室内饲养到成虫后作为试验虫源。高密度种群放入80 cm×80 cm×80 cm的笼中群体饲养,每笼50~60头;低密度种群放入10 cm×10 cm×20 cm的笼中进行单头饲养。以小麦苗等禾草饲喂,并辅以麦麸补充营养。蝗虫达到成虫期雌雄分别饲养,不进行交配。温度控制在26~28 ℃,光周期L∥D=14 h∥10 h,相对湿度为50%~60%。飞行测试的各日龄成虫选取体型相当(以体长相等为主)、体质量相近的个体作为试虫。高密度种群采集地点为 41°47.902′ N,111°57.799′ E,低密度种群为40°47.156′ N,103°12.622′ E。
1.2飞行能力测试 飞行测试利用中国农业科学院植物保护研究所自行研制的昆虫飞行数据微机采集系统[11]。为了适应亚洲小车蝗个体体质量较大的特点,制作了适宜亚洲小车蝗体质量的悬臂和相应配重。试虫固定参照张龙和李洪海[12]的方法并进行相应改进,用细铜丝线和502胶将试虫固定于飞行磨悬臂上。试虫吊好后打开微机进行飞行数据采集。吊飞12 h后停机,用微机将测试结果输出,采集系统为10通道,每次悬吊10头试虫,共测40头,其中雌、雄各20头。吊飞室内温度保持在26~28 ℃,相对湿度50%~60%;测试时间为20:00至次日08:00;吊飞期间保持黑暗条件。测定的主要飞行能力参数为飞行距离、飞行时间、飞行速度和最大飞行速度。
1.3数据统计分析 所有数据分析均在DPS统计分析软件上进行[13]。采用t检验分析不同密度条件的亚洲小车蝗成虫的飞行能力;对不同日龄和性别的飞行能力进行方差分析,Duncan法进行多重比较。
2 结果与分析
2.1亚洲小车蝗飞行能力与成虫日龄的关系 成虫的飞行能力随成虫日龄增加而发生变化。初羽化及1日龄成虫由于虫体较软、飞行肌发育不完全,飞行能力弱,不具备远距离飞行的能力;2日龄后成虫飞行能力逐渐增强。因此,亚洲小车蝗飞行能力从4日龄开始测定。
不同日龄的亚洲小车蝗成虫飞行能力存在显著差异。在12 h的飞行测试中,4日龄成虫具有了一定的飞行能力,单头平均累计飞行时间可达0.2 h,平均累计飞行距离2 km左右;7日龄成虫的飞行能力开始增强;13日龄达到最高峰,平均累计飞行时间可达1.42 h,累计飞行距离近15 km;20日龄开始下降,但仍有较强的飞行能力。从结果可以看出,成虫的平均飞行速度也随着日龄增长而逐渐增大,4、7日龄成虫平均飞行速度明显低于10、13、20日龄,10日龄后成虫飞行速度变化不大(表1)。
亚洲小车蝗成虫不同性别比较结果表明(表1、表2),雌虫飞行能力(飞行距离和飞行时间)略高于雄虫,不同日龄雌雄成虫飞行能力的变化趋势基本一致。雌雄之间的飞行参数均相近,即性别间的飞行能力差异不大。雄虫4日龄飞行能力最弱,13日龄飞行能力达到最大。

表1 亚洲小车蝗雌虫飞行能力与虫龄关系Table 1 Relationship between the age and flight ability of female Oedaleus asiaticus
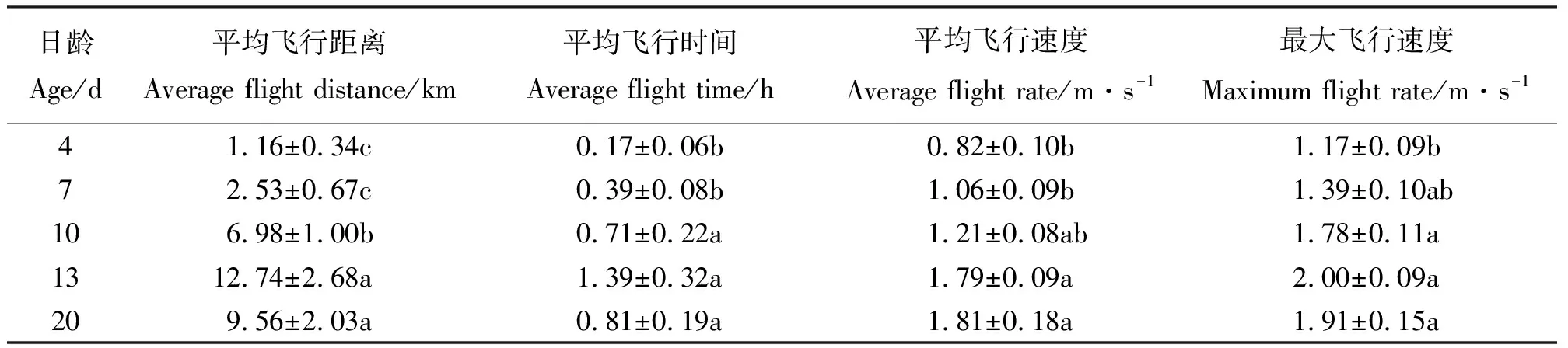
表2 亚洲小车蝗雄虫飞行能力与虫龄关系Table 2 Relationship between the age and flight ability of male Oedaleus asiaticus
2.2亚洲小车蝗飞行能力与种群密度的关系 12 h的吊飞测试结果表明(图1、图2),种群密度对亚洲小车蝗飞行能力有显著影响,即同日龄的亚洲小车蝗高密度种群飞行能力显著高于低密度种群,雌雄虫测试结果一致。
高密度种群雌虫最大飞行距离可达14 km,雄虫可达10 km,在13日龄飞行能力最强。平均飞行时间雌虫近1.5 h,雄虫近1.2 h,表现出较强的飞行能力。低密度种群各日龄也有一定的飞行能力,雌雄之间差异不明显。13日龄飞行距离最大,这点与高密度种群相同,各日龄的单头累计飞行距离大多在3 km左右(图1、图2)。结果表明,低密度种群亚洲小车蝗的飞行能力不强。
3 讨论与结论
昆虫的迁飞行为一直是国内外昆虫研究领域中的热点[14-15],应用飞行磨来研究昆虫的飞行行为虽不能完全模拟或表达昆虫飞行的自然状况,但迄今为止在昆虫的生理、生态研究中仍是被普遍认可和应用的一种实验工具[16]。许多昆虫学家通过飞行磨吊飞测试研究昆虫的飞行行为[17-21],张龙等[22]利用飞行磨研究过两型东亚飞蝗(Locustamigratoria)飞行能力的差异;刘辉等[23]对群居型与散居型东亚飞蝗飞行能力进行了比较研究。目前对亚洲小车蝗这一重大草原害虫的飞行能力及迁移生物学等方面还处于空白状态,有许多基础科学问题尚不明确。
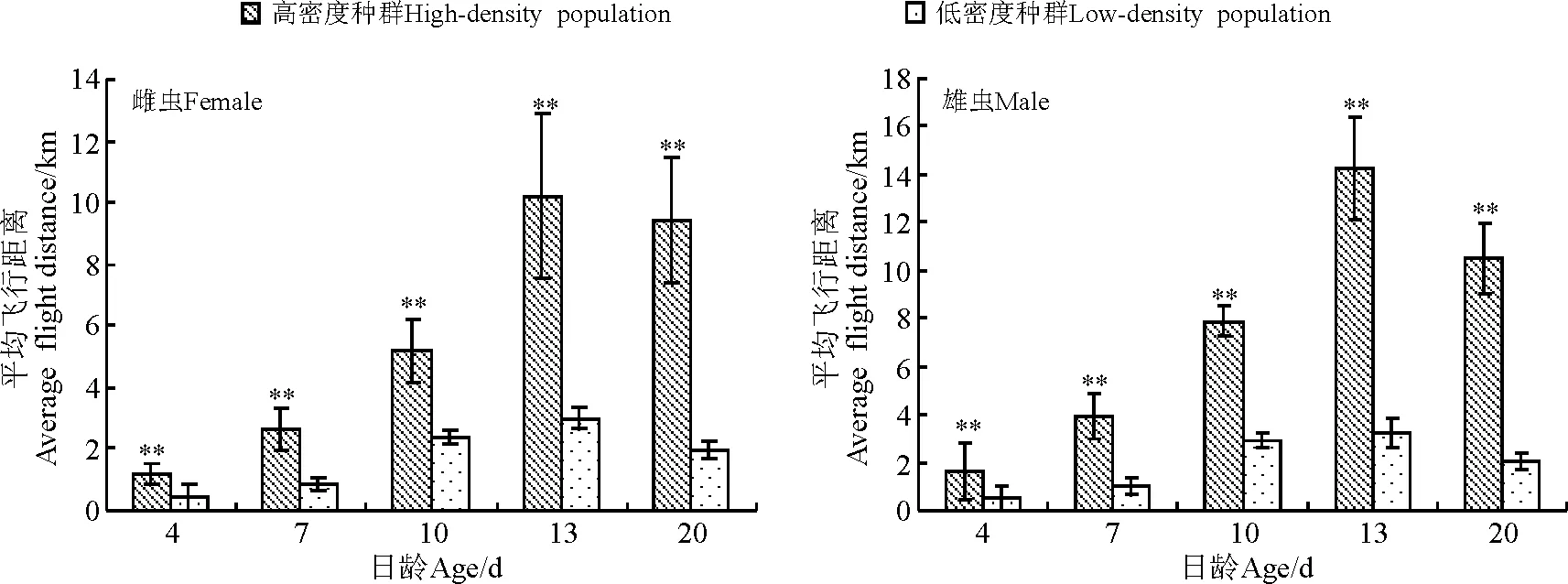
图1 不同种群密度的亚洲小车蝗虫的平均飞行距离Fig.1 Average flight distance of Oedaleus asiaticus in different population densities
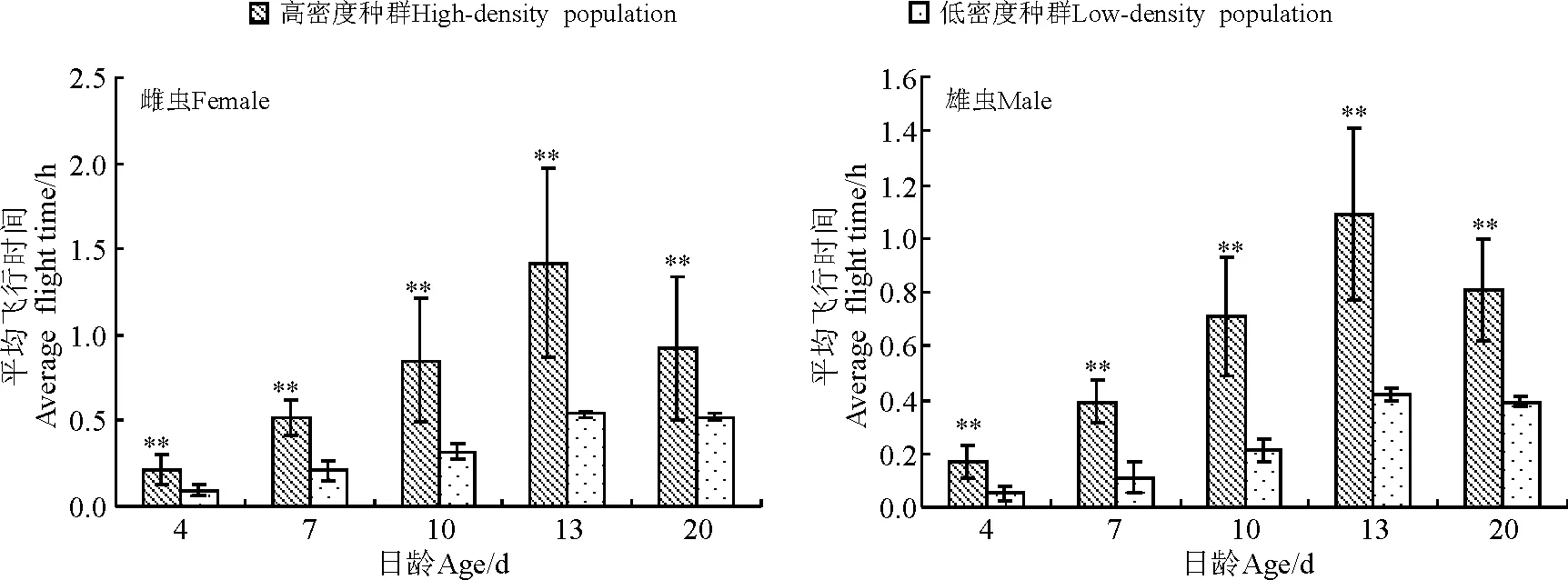
图2 不同种群密度的亚洲小车蝗虫的平均飞行时间Fig.2 Average flight time of Oedaleus asiaticus in different population densities
种群密度是诱导昆虫迁飞行为发生的重要环境因子之一[24]。种群密度同环境因子(光周期、温度)一样,都是影响昆虫迁飞行为发生的重要因子[25],虽然遗传因子对昆虫的飞行能力有一定影响,但是环境因子的作用更为重要,尤其种群密度对昆虫的飞行能力有直接影响[26]。研究表明,种群密度增加对马铃薯甲虫(Leptinotarsadecemlineata)越冬代成虫飞行具有明显的刺激作用[27];东亚飞蝗(L.migratoria)生态型转变与种群密度有显著的相关性,而且种群密度高低可影响其从群居型向散居型转化的速度[28];幼虫密度可影响非洲粘虫(Spodopteraexempta)变型,即高密度条件下产生的群居型、低密度产生的散居型以及居于两者之间的中间型[29]。
亚洲小车蝗属于土蝗,虽然近年来很多人观察到它的迁飞行为,但是对其迁飞习性还没有确定,它是否具有群居型与散居型的划分也不明确,高密度和低密度种群间形态上没有明显的差异。本研究表明,不同种群密度对亚洲小车蝗的飞行能力有显著影响,亚洲小车蝗高密度种群平均累计飞行距离近15 km,累计飞行时间可达1.42 h,而低密度种群成虫累计飞行距离大多在3 km左右,累计飞行时间可达0.31 h。亚洲小车蝗高密度种群的飞行能力显著高于低密度种群。东亚飞蝗飞行能力也有类似表现,13日龄的散居型东亚飞蝗平均累计飞行距离和时间比群居型分别低17~18倍[23]。高密度与食物数量、质量下降常常并存发生,而且食物资源缺乏能够诱使种群迁飞行为的发生。以上结果说明,亚洲小车蝗可能具有群居型和散居型的分化,而且亚洲小车蝗聚集爆发时具有迁飞的可能。东亚飞蝗的累积飞行能力超过35 km[17],本研究显示,亚洲小车蝗的飞行能力较东亚飞蝗低很多,可能受所取样本数及室内环境条件限制,因此,需要结合田间试验进行深入研究。
目前,学者们总结了国内外蝗虫研究状况和近期新进展,但是蝗虫的飞行能力以及迁飞等相关研究工作涉及很少[30]。我国仅对东亚飞蝗振翅频率、飞行能力等特性开展过一些研究,亚洲小车蝗飞行能力及相关机制的研究尚处于空白状态,亟待加强,蝗虫出现散居型向群居型转变和群居型的大规模迁飞是其猖獗危害的重要特点。因此,如何控制群居型群体的形成是控制蝗灾发生的关键。本研究初步明确了亚洲小车蝗飞行能力及其与种群密度的关系,表明其可能具有群居型和散居型的分化,这对于开展相关研究有一定的借鉴意义。
[1] 刘玲,郭安红.2004年内蒙古草原蝗虫大发生的气象生态条件分析[J].气象,2004,30(11):55-57.
[2] 潘建梅.内蒙古草原蝗虫发生原因及防治对策[J].中国草地,2002,24(6):66-69.
[3] Kang L,Chen Y L.Dynamics of grasshopper communities under different grazing intensities in Inner Mongolia steppes[J].Entomologia Sinica,1995,2:265-281.
[4] 蒋湘,买买提明,张龙.夜间迁飞的亚洲小车蝗[J].草地学报,2003,11(1):75-77.
[5] Cheke R A.A migrant pest in the Sahel:the Senegalese grasshopperOedaleusenegalensis[J].Philosophical Transactions of the Royal Society of London.Series B Biological Sciences,1990,328(1251):539-553.
[6] Riley J R,Reynolds D R.Nocturnal grasshopper migration in West Africa:Ttransport and concentration by wind,and the implications for air to air control[J].Philosophical Translations of the Royal Society of London.Series B. Biological Sciences,1990,328(1251):655-672.
[7] 牛虎力,周强.草原蝗虫的红外灯光诱集实验初报[A].农业生物灾害预防与控制研究[C].杭州:中国植物保护学会第九届会员代表大会暨2005年学术年会,2005.
[8] 乔峰.蝗灾北移的主力军——亚洲小车蝗[J].人与生物圈,2005(3):23-25.
[9] 关敬群,魏增柱.亚洲小车蝗(OedaleusasiaticusB.-bienko)食量测定[J].昆虫知识,1989,26(1):8-10.
[10] 李广.亚洲小车蝗为害草场损失估计分析的研究[D].北京:中国农业科学院,2007.
[11] 程登发,田品,孙京瑞,等.适用于蚜虫等微小昆虫的飞行磨系统[J].昆虫学报,1997,40(增刊):172-179.
[12] 张龙,李洪海.虫口密度和龄期对东亚飞蝗群居型向散居型转变的影响[J].植保技术与推广,2002,22(4):30-35.
[13] 唐启义,冯明光.实用统计分析及其计算机处理平台[M].北京:科学出版社,2002:34-130.
[14] Rankin M A,Burchsted J C A.The cost of migration in insects[J].Annual Review of Entomology,1992,37:533-559.
[15] 江幸福,罗礼智.昆虫迁飞的调控基础及展望[J].生态学报,2008,28(6):2835-2842.
[16] Dingle H.Adaptive variation in the evolution of insect migration[A].In:Rabb R L,Kennedy G G.Movement of Highly Mobile Insects:Concepts and Methodology in Research[M].Raleigh, North Carolina:North Carolina State University Press,1979:64-87.
[17] 罗礼智,李光博,胡毅.粘虫飞行与产卵的关系[J].昆虫学报,1995,38(3):284-289.
[18] 曹雅忠,黄葵,李光博.空气相对湿度对粘虫飞翔活动的影响[J].植物保护学报,1995,2(2):134-138.
[19] 江幸福,蔡彬,罗礼智,等.温、湿度综合效应对枯虫蛾飞行能力的影响[J].生态学报,2003,23(4):738-743.
[20] 尹蛟,封洪强,程登发,等.粘虫成虫在气流场中飞行行为的观察研究[J].昆虫学报,2003,46(6):732-738.
[21] 贾佩华,曹雅忠.小地老虎成虫飞翔活动[J].昆虫学报,1992,35(1):59-65.
[22] 张龙,严毓华,李光博,等.蝗虫微孢子虫病对东亚飞蝗飞行能力的影响[J].草地学报,1995,3(4):324-347.
[23] 刘辉,李克斌,尹姣,等.群居型与散居型东亚飞蝗飞行能力的比较研究[J].植物保护,2007,33(2):34-37.
[24] Tammaru T,Ruohomki K,Montola M.Crowding-induced plasticity inEpirritaautumnata(Lepidoptera:Geometridae):Weak evidence of specific modifications in reaction norms[J].Oikos,2000,90:171-181.
[25] Shimizu T,Masaki S.Injury causes microptery in the ground cricket,Dianemobiusfascipes[J].Journal of Insect Physiology,1993,39:1021-1027.
[26] Knor I B,Bashev A N,Alekseev A A,etal.Effect of population density on the dynamics of the beet webwormLoxostegesticticalisL.(Lepidoptera:Pyralidae)[J].Entomology Review,1993,72:117-124.
[27] 郭利娜,郭文超,刘曼双,等.拥挤度对马铃薯甲虫飞行能力的影响[J].新疆农业科学,2011,48(2):320-327.
[28] 张龙,李洪海.虫口密度和龄期对东亚飞蝗群居型向散居型转变的影响[J].植保技术与推广,2002,22(4):3-5.
[29] Faure J C.Phase variation in the armyworm,Laphygmaexempta(Walk.)[M].Scientific Bulletin 234.Pretoria:Department of Agriculture and Forestry of south Africa,1943.
[30] 王杰臣,倪绍祥.国内外蝗虫研究发展动向初探[J].干旱区研究,2001,18(3):36-40.
