青海云杉半同胞家系苗期遗传变异及选择1)
2012-03-05刘林英张宋智马建伟安三平
刘林英 蒋 明 张宋智 马建伟 安三平
(甘肃小陇山林业科学研究所,天水,741022)
王军辉
(中国林业科学研究院林业研究所(国家林业局林木培育重点实验室))
青海云杉(Picea crassifolia)是我国青藏高原东北边缘特有树种,是我国北方地区造林和城市绿化的重要树种,主要分布于青海、甘肃、宁夏、内蒙古等地[1-2]。长期以来青海云杉以自然采种繁殖为主,育苗周期长、生长缓慢、优良基因资源贫乏而影响该树种的发展[3-4]。近年来,国内外在云杉属树种的强化育苗方面进行了一系列的研究,取得了许多研究成果,使青海云杉苗期的生长周期缩短一半以上[5-6]。但对其优良种源、家系的苗期测定及选择还未有更深入地研究。为了提高青海云杉的林地生产力,需要选择出遗传增益大,速生优良的家系进行生产推广。在对青海云杉基因资源调查收集的基础上,笔者于2006年在青海省大通县东峡林场,采集青海云杉优树67 个家系种子。2007年至2009年,采取强化育苗技术措施进行培育,通过对保留数量在50 株以上的51 个家系苗期生长性状的测定,比较家系间苗期形态指标的差异,了解青海云杉优树家系的苗期性状变异规律,并对青海云杉优良家系进行苗期评定和选择,为青海云杉高世代育种奠定基础。
1 试验地概况
试验设在甘肃省天水市小陇山林业科学研究所苗圃,海拔1 160 m。年降水量500 ~700 mm,年蒸发量1 290.0 mm,年平均气温12 ℃,极端高温39℃,极端低温-19 ℃。无霜期约200 d。
2 材料与方法
材料来源与试验布置:2006年在青海省大通县东峡林场青海云杉初级无性系种子园中采收了67个家系种子。2007年采用塑料育苗袋(8 cm×14 cm)育苗,完全随机区组排列,每家系播种200 袋10行小区,播种前所有家系种子用高锰酸钾溶液消毒后催芽,每家系50%露白时播种,每袋3 粒。在苗木生长期的4月至8月连续2 a,采用功率36 W 的日光灯夜间补光,每天从22:00 至次日2:00 对苗木补光4 h,光照强度1 ~6 μmol/(m2·s)。2009年3月将苗木移植于(15 cm×18 cm)容器袋中培育,未进行补光处理。
生长指标测定方法:2009年秋末,对保存数量在50 株以上的3年生51 个家系苗随机抽取50 株,每次10 株5 次重复。测量每个单株的苗高和地径等生长指标。然后每个家系选取15 株标准木,测定每个单株的生物量。每小区家系里选取15 株苗木,洗净、晾干,测量每株苗木的苗高、地径、新梢长、侧枝数、侧枝长和根系生长指标后,将苗木沿根茎处剪断,分别挂上标签,装入纸袋,进行烘干。用烘箱在80 ℃的条件下烘干至恒质量后分别称质量,记录每株苗木的茎叶及根系干质量。计算高径比、径根比和质量指标。其中,高径比=苗高/地径;径根比=茎叶干质量/根干质量;质量指标=苗木总干质量/[(苗高/地径)+(茎干质量/根干质量)]。
统计分析方法:对数据采用Excel 软件和DPS10.15 进行数据处理、方差和相关分析。计算家系遗传力、遗传增益和变异系数[7-11]。
3 结果与分析
3.1 青海云杉家系生长性状的变异
青海云杉苗高、地径、新稍长、分枝角度、侧枝数、侧枝长、侧根数、侧根长家系间存在极显著差异(P<0.01),主根长、径根比和质量指数存在显著差异(P<0.05),高径比差异不显著(表1)。遗传变异系数反应性状在群体中遗传变异的幅度,用遗传变异系数可以反映遗传因素导致性状相对变异的程度,遗传变异系数大,说明家系对该性状的可利用的改良潜力较大,对其改良可取得较好的效果。苗高、地径、新梢长、分枝角度、侧枝数、侧枝长、主根长、侧根数、侧根长、径根比和质量指标的遗传变异系数较大,均在11.50%以上,说明青海云杉家系具有丰富的遗传变异,开展家系选择具有很大的潜力。
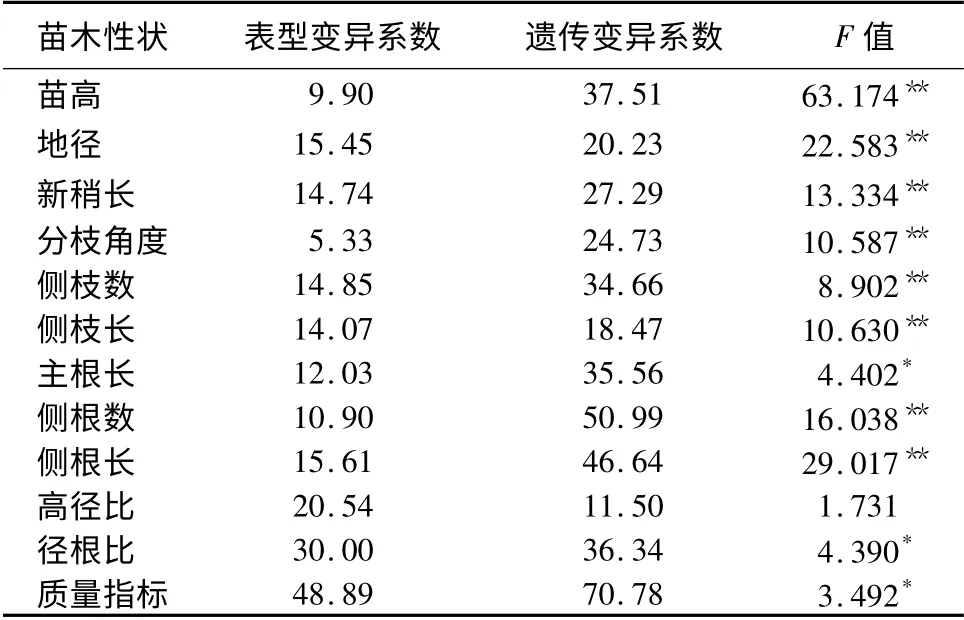
表1 青海云杉家系苗期各性状的变异
3.2 青海云杉家系生长性状的遗传参数估算
青海云杉苗高、地径、新梢长、侧枝数、侧枝长、侧根长、高径比、径根比和质量指标的家系遗传力均较大,均在0.80 以上,其中地径的遗传力最大,为0.993。主根长、分枝角度和侧根数的遗传力较低,其中主根长的遗传力最低,为0.239,主根生长受到容器的限制,表现出低度遗传(表2)。方差分量表明,家系间青海云杉实生苗各性状的变异较大。因此,各家系形状间有丰富的变异,这些变异是优良家系选择的基础。
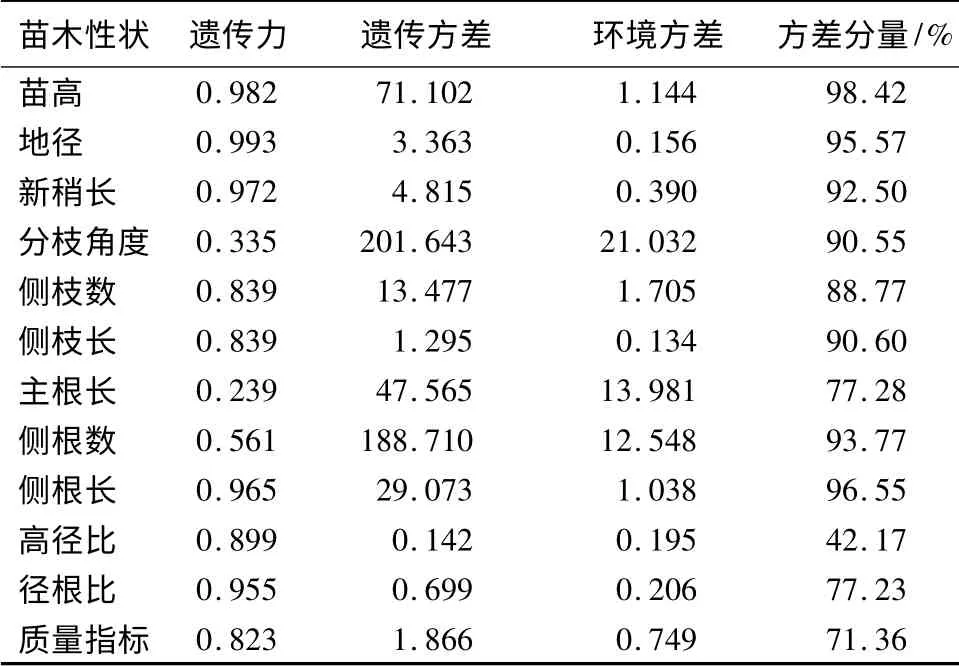
表2 青海云杉家系生长性状遗传参数分析
3.3 青海云杉家系间各生长性状的相关分析
青海云杉苗期各性状相关分析表明,地上指标与根系生长存在一定的相关性。苗高与地径、新梢长、侧枝数、侧根长呈极显著的正相关,相关系数分别为0.76、0.69、0.66、0.51;表明苗木高度生长量受影响光合面积大小的新梢长、侧枝数以及根系大小的控制,光合面积和根系吸收面积越大,苗高生长量也较大,同时从新梢长度也可判断出苗木的总体高度。地径与新梢长、侧枝数、主根长、侧根数、侧根长呈极显著正相关,相关系数分别为0.77、0.63、0.56、0.45、0.66。这种相关性表明,苗木地径生长量主要受光合面积、根系营养吸收范围影响,苗木根系越发达,地径越大,反过来可以说地径大的苗木,其新梢生长量也会较大。新梢长与侧枝数、主根长、侧根长,侧枝数与主根长、侧根长,主根长与侧根数均呈极显著的正相关,相关系数分别0.55、0.49,0.40、0.48、0.36、0.51;苗高与侧根数,新梢长与侧根数,侧枝数与侧根数均呈显著正相关,相关系数分别0.29、0.34、0.32;此相关结果说明,苗木形态性状之间具有紧密的联系,苗木地上部分的生长量受地下部分影响,同时地上部分的光合产物促进了地下部分的生长和吸收功能的增强。径根比与苗高、新梢长、侧枝数、侧根数均呈负相关,为-0. 07、-0.22、-0.11、-0.23,径根比与地径、主根长、侧根长呈极显著负相关,为-0.41、-0.37、-0.44(表3)。表明径根比越大,根系越差。其原因一方面是青海云杉生物学特性的表现,另一方面可能是大棚补光育苗,环境的改善,使苗木地上部分得到了充分的生长,而地下部分受容器的控制而造成。
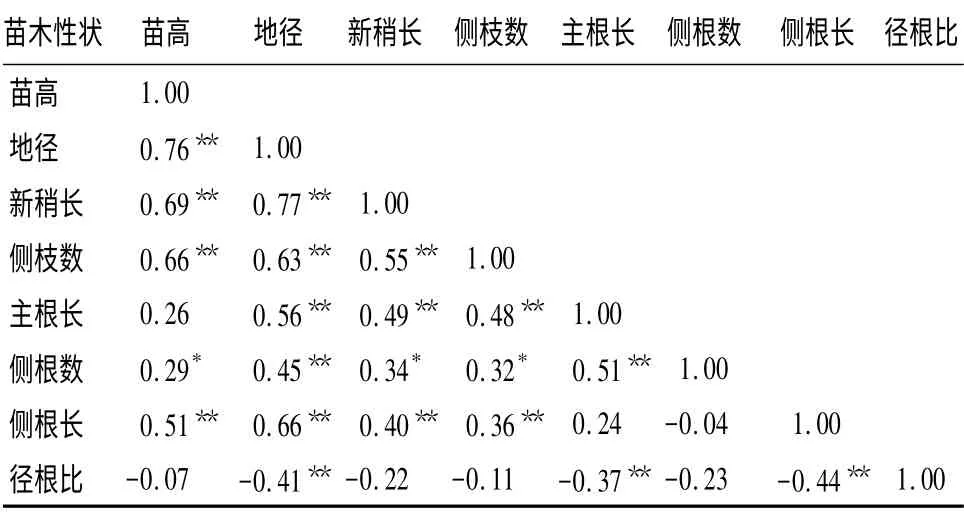
表3 青海云杉苗期生长性状与根系性状的相关分析
3.4 不同家系性状的综合分析及选择
结合方差分析及相关性分析结果,3年生苗的苗高、地径、新梢长、侧枝数、侧根长的遗传变异系数、遗传相关系数、遗传力均较高,说明该群体的遗传潜力大。各性状的表型变异系数和环境变异系数比遗传变异系数低,说明性状主要受遗传控制,所以在苗期进行选择时,可以根据其中一个性状苗高为标准对青海云杉优良家系初选。51 个家系的平均苗高为22.48 cm,标准差为4.95,苗高大于1.5 倍标准差的家系有311#、312#、405#、109#,苗高大于1倍标准差的家系有311#、312#、405#、109#、408#、410#、305#、306#,它们对应的多项生长指标都表现优良,特别是311#家系高生长较快,平均值为35.62 cm,其次是312#(34.23 cm)、405#(32.38 cm)、109#(31.23 cm),这4 个生长较快的家系的平均值为3 3. 37 cm,4 个生长快的家系超过群体平均值的48.44%,生长最快的311 号家系超过群体平均值的58.45%。
对供试的51 个青海云杉家系,在中选率为15.69%的水平下进行计算,初选优良家系8 个,分别为311#、312#、405#、109#、408#、410#、305#、306#。所选家系在生长和根系性状方面有很大提高,平均苗高30.81 cm,相比原群体提高37.05%;平均地径0.86 cm,增加24.63%,平均侧枝数12.73 cm,增加20.39%,其他指标均有较大地提高。综合选择的8个青海云杉优良家系能获得较高的遗传增益,苗高、地径、新梢长、侧枝数、主根长、侧根数、侧根长、径根比9 个性状的遗传增益分别为36.39%、21.37%、2 2.12、%、17.12%、1.98%、6.33%、26.45%、-8.70%。其中苗高的遗传增益最大,说明选择效果显著。
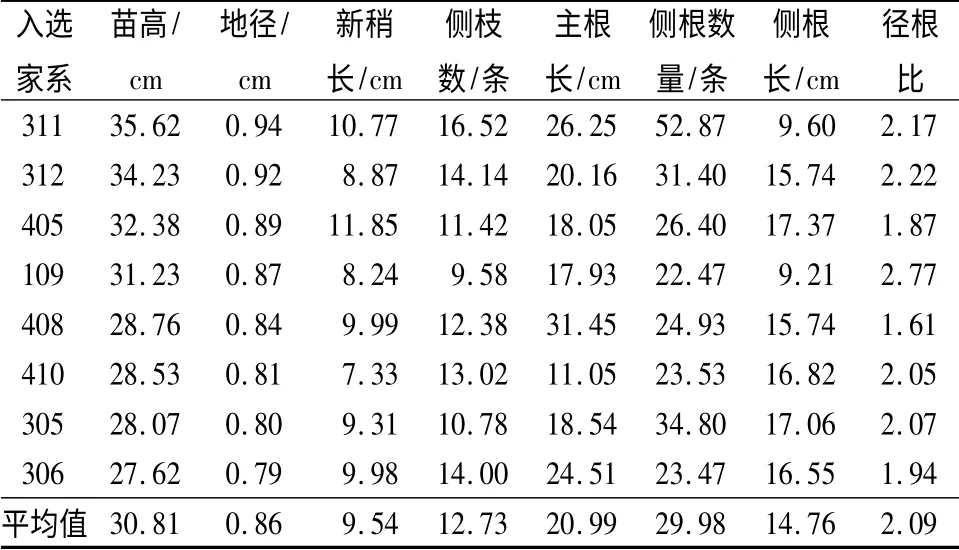
表4 青海云杉入选优良家系的平均值
4 结论与讨论
苗高和地径是反映苗木质量最直观的指标[12]。苗期生长测定结果表明,青海云杉家系间在苗高、地径生长方面存在极显著差异,苗高家系变幅为17.69 ~35.62 cm,地径家系变幅为0.48 ~0.94 cm。遗传参数结果表明,苗高遗传力为0.982,遗传变异系数为37.51%;地径遗传力为0.993,遗传变异数为20.23%。因此,以苗高和地径为指标进行选择,都会取得较好的遗传改良效果。相对地径而言,用苗高性状作为主要选择指标,效果更佳。
新梢长、侧枝数、侧枝长、分枝角度是苗木地上部分生长的主要特征。通过测定青海云杉不同家系的地上生长性状,发现新梢长,侧枝数和侧枝长家系间存在极显著差异,遗传力较高,呈强度遗传,同时侧枝数、侧枝长和分枝角的大小与地径等性状存在显著的相关关系,因此在育种过程中侧枝的性状也是不能忽视的。
根系与地上部分作为植株生长过程的2 大功能器官,是相互依存竞争的统一体[11],根系的生长发育状况与地上部的生长发育状况息息相关[13-14]。由于根系是反映苗木耐干旱,吸收养分等的重要指标,它与地上部是一个有机整体[15]。对青海云杉家系苗期根系生长状况分析表明,侧根数和侧根长家系间的差异极显著,主根长家系间差异显著。遗传力反映了亲代把性状传递给子代的能力,是遗传育种中一个重要参数[12]。除主根长外,其它根系性状遗传力较大,呈强度遗传,其中侧根长遗传力最大,为0.965,主根长遗传力最低,为0.239,呈低强度遗传。理论上苗木根系应该是呈强度遗传,由于试验采取容器育苗方式,苗木主根生长受到很大限制,所以造成主根长的遗传力低下,但促进了侧根的发展。
通过相关性分析,青海云杉家系苗高、地径、新梢长、侧枝数之间存在极显著的正相关,侧根数与地径、新梢长、侧枝数均呈显著或极显著正相关,而与根系生长指标之间相关性较小,侧根数与侧根长、径根比之间均呈负相关和极显著负相关,说明侧根数越多,苗期生长越弱,径根比越大,反映了根系生长越差,进一步说明根系参数也是评价青海云杉苗期生长,遗传育种的重要指标。
综合生长形态指标并结合苗高分析结果,初步选定311#、312#、405#、109#、408#、410#、305#和306#为优良家系,它们的多项生长指标表现优良,遗传增益明显,具有生产使用价值。本研究只是苗期的初步选择,结果将会在今后的家系试验林中开展跟踪测定进行检测,以期为青海云杉家系选择提供科学合理的参考。
[1] 刘兴聪.青海云杉[M].兰州:兰州大学出版社,1992.
[2] 王豁然,江泽平,傅紫岐. 林木引种驯化与森林可持续经营[M].北京:中国环境科学出版社,1998:165-171.
[3] 马常耕.世界云杉无性系林业发展现状[J]. 世界林业研究,1993(6):24-31.
[4] Luo Jianxun,Zhang Xiaolu,Gu wanchun. Biogeographic differences in cone,needle and seed morphology among native Picea asperata populations in Western China[J]. Forestry Studies in China,2005,7(2):1-6.
[5] 张守攻,王军辉,刘娇妹,等.青海云杉强化育苗技术研究[J].西北农林科技大学学报:自然科学版,2005,33(5):33-38.
[6] 张华丽,张金凤,王军辉,等. 针叶树补光育苗技术研究进展[J].西北林学院学报,2005,20(1):107-111.
[7] 沈熙环.林木育种学[M].北京:中国林林业出版社,1990.
[8] 柯潮榕.马尾松半同胞子代测定及速生优良家系和个体选择[J].福建林业科技,2008,35(2):25-29.
[9] 邓继峰,张含国,张磊,等.17 种杂种落叶松遗传变异及优良家系选择[J].东北林业大学学报,2010,38(1):8-11.
[10] 张含国,张磊,李雪峰,等. 长白落叶松优树选择效果的评价[J].东北林业大学学报,2007,35(9):1-3,11.
[11] 赵秉强,张福锁,李增嘉,等. 套作夏玉米根系数量与活性的空间分布及变化规律[J].植物营养与肥料学报,2003,9(1):81-86.
[12] 董章凯,邢世岩,王亚明,等. 麻栎半同胞家系苗期特性分析[J].东北林业大学学报,2011,39(4):27-28,36.
[13] 王国良,罗建勋,文吉富,等. 马尾松种子园半同胞家系子代苗期性状遗传变异[J].四川林业科技,2009,30(3):18-21,17.
[14] 姜静,杨光,祝泽兵,等.白桦强化种子园优良家系选择[J].东北林业大学学报,2011,39(1):1-4.
[15] 张锁,樊军锋,刘永红,等. 油松自由授粉子代抗逆性性状遗传分析与初选[J].西北林学院学报,2011,26(3):93-98.
