浮筏式养殖中海带个体生产力与夹苗位置之间的相关性分析
2012-01-12袁著涛刘升平
任 伟, 袁著涛, 刘升平
(青岛农业大学, 山东 青岛 266109)
浮筏式养殖中海带个体生产力与夹苗位置之间的相关性分析
任 伟, 袁著涛, 刘升平
(青岛农业大学, 山东 青岛 266109)
在浮筏式养殖中, 海带的个体生产力与夹苗位置密切相关, 上水层的海带长势较好。在育种中按照生产性状优劣的方法选种时, 可被选种的个体仅占总个体的 10%左右。本文采用同一对配子体无性繁殖系培育的克隆苗为实验材料, 进行了个体生产力与夹苗位置之间的相关性分析。实验结果表明:在长度、宽度、厚度、鲜质量、干质量等各项指标中, 干质量最能体现海带的个体生产力, 得到方程式:y=-0.010x+0.387(R=0.933,P<0.01), 该方程式表达了夹苗位置对海带个体生产力的影响。在海带育种选种时, 利用公式y=0.010(n-1)+x, 可以通过各个夹苗位置的实测值计算出最佳夹苗位置的期望值,消除因环境因素对海带个体生产力带来的影响, 使实验结果之间有了可比性, 能避免选种中的漏选或误选。
海带; 浮筏式养殖; 个体生产力; 夹苗位置; 育种
在海带育种工作中, 海带选种的依据包括叶片长度、宽度、厚度、个体质量、鲜干比及色泽等, 上述指标决定了海带的个体生产力。影响海带个体生产力的主要因素是基因型(品种)和环境条件, 在浮筏式养殖中, 经过长期的生产实践, 每个海域都找到了最佳养殖技术指标, 此时, 夹苗位置是影响海带个体生产力的最主要的环境因素。
在浮筏式养殖海带中, “平养法”与“垂养法”比较,一定程度上解决了上层海带对下层海带的遮光问题[1], 但由于重力的因素, 整绳海带并非处于同一水平位置, 导致了上层海带长势较好, 而这个比例约占整绳海带的 10%左右,生产中这个问题可以通过加小浮子得到部分解决。由于生产中追求的是群体生产力, 所以这个问题并非必须解决, 但是海带育种选种时就只能从上层 10%海带中选择, 造成了选种时的浪费及对可能存在的优良海带种质资源的丢失:一方面, 上层个体生产力较大的海带可能只是表现型的结果, 基因型并不一定好, 可能造成误选; 另一方面, 下层的海带由于受环境制约[2-4], 生长潜力并没有得到体现, 而这些海带中可能存在基因型优良的海带, 造成漏选。
在通常情况下, 浮筏式养殖海带的个体生产力由夹苗位置及基因型决定。为了消除环境因素对海带生产性状表达的影响, 做出夹苗位置对海带个体生产力的相关性分析是必要的。海带无性繁殖系具有遗传的专一性和发育全能性的特点[5], 同一对配子体无性繁殖系培育的海带克隆苗可以解决基因型不同的问题, 利用基因型完全一致的海带苗种进行养殖实验, 对苗绳上所有海带的生长结果做出统计分析, 得出表达海带个体生产力的各项指标与夹苗位置的相关方程, 把实测值转化为同一夹苗位置下的期望值, 使之具有可比性, 就可以准确地比较基因型的优劣, 避免选种时的漏选或误选。
1 材料与方法
1.1 实验海域条件
实验海域位于山东半岛东部荣成市俚岛湾养殖区, 由于夏季受黄海冷水团的影响, 表层水温超过23℃的时间较短, 因此在温度上非常适合海带养殖。又因实验海区是渤海发源的沿岸流的影响区域[6],给海带生长带来了的丰富营养盐。自1952年海带筏式养殖实验成功后, 实验海域就成为中国主要的海带养殖基地[7]。
1.2 材料与方法
1.2.1 材料
苗种:实验用苗种采用本实验室保存的一对海带配子体培育的克隆苗[8-9]。
苗绳:长度2 m, 直径约15 mm的聚乙烯、红棕丝混纺苗绳。
1.2.2 方法
夹苗:2010年12月17日进行海带夹苗, 2011年3月9日, 海带平均长度为60 cm时, 选择规格相同的海带苗二次夹苗。
养殖管理:海带苗间距为 7~8 cm, 把两根苗绳连接, 将苗绳两端用吊绳固定在海带筏架上, 筏间距约为4 m。由于悬浮在海水中的泥沙和杂藻孢子可能影响海带的生长, 并可能引起病害, 所以每隔一周冲洗一次海带叶片表面的浮泥[10]。
烘干处理:海带收获后, 将海带置于 60℃的烘干箱中烘干24 h, 然后称干质量。
1.3 数据测量
在离海带叶片基部40 cm处打孔, 记录原始数据,以后每隔10天采集一次数据。表达个体生产力各项指标:海带长度L(cm)、宽度W(cm)、厚度H(mm)、个体鲜质量G1(kg)、个体干质量G2(kg); 其他因子数据:水温T(℃)、赛氏透明度DS(m)、海带夹苗位置n、海带生长期d(d)。各项目的测定均采用标准方法[11]。
各个海带苗绳的实验结果用Microsoft Excel按照时间顺序整理, 并将其转化为数据集。由于海带处于水文指标相同的同一海区, 其他环境因素可以认为相同。将各根海带苗绳的实验结果进行算术平均,得到平均值。
1.4 统计分析
实验数据分析在SPSS 17.0中进行, 对实验数据用 one-way ANOVA 进行分析, 然后对数据方差不齐的进行再处理, 待数据合理后进行显著性及相关性分析。
2 结果与分析
2.1 生长期、夹苗位置对海带长度的影响
从3月19日到海带收获, 其长度生长曲线(图1)可以表示为:
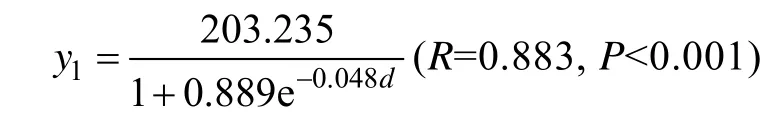
式中y1为长度,d为生长期。
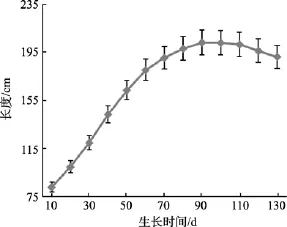
图1 长度生长曲线Fig.1 Corelation between the length and time
表1反映了海带长度在不同时间与夹苗位置的相关系数, 3月份海带处于凹凸期, 叶片较小且在水流作用下悬浮, 相互间影响小, 随着海带生长, 叶片变大, 相互遮光, 并在重力作用下下垂, 海带长度与夹苗位置的相关性越来越高。到6月中下旬, 水温上升(图2), 上水层海带梢部开始腐烂, 其生长速度小于腐烂速度, 海带的相关性又趋于降低。到7月上旬海带选种时, 海带长度与夹苗位置的方程为:
y1=-1.555n+216.81(R=0.853,P<0.01)
式中y1代表长度,n为夹苗位置。此时, 海带长度的实测值, 与最佳水层的期望值的关系为:y′1=1.555(n-1)+x1。
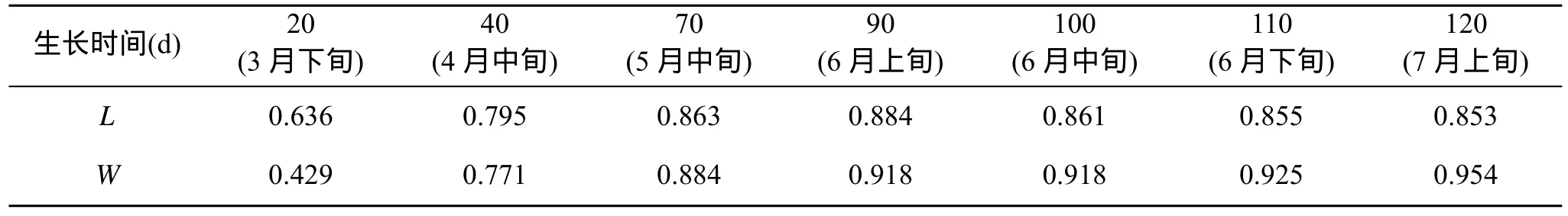
表1 海带的长(L)、宽(W)在不同生长阶段与夹苗位置的相关性比较Tab. 1 Correlation between the length(L) and width(W) of Laminaria japonica at different growth stages and clip seedlings position
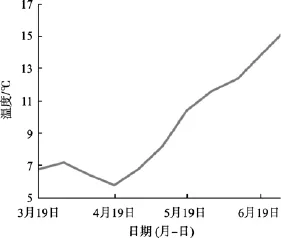
图2 实验海区水温变化曲线Fig. 2 Variation of temperature in the sea
2.2 生长期、夹苗位置对海带宽度的影响
海带宽度是估算海带生产力的另一个指标, 其宽度生长曲线(图3)可以表示为:
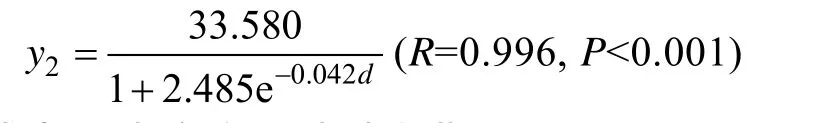
式中y2为宽度,d为生长期。
夹苗位置对海带宽度的影响随海带个体增大,其相关性越来越好(表1)。到7月上旬选种时达到了0.954, 其方程式为:
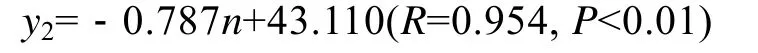
式中y2为宽度,n为夹苗位置。此时, 海带宽度的实测值, 与最佳水层的期望值的关系为:y′2=0.787(n-1)+x2。
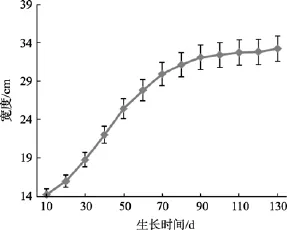
图3 宽度生长曲线Fig. 3 Corelation between the width and time
2.3 生长期、夹苗位置对海带厚度的影响
厚度的生长曲线(图4)可以表示为:
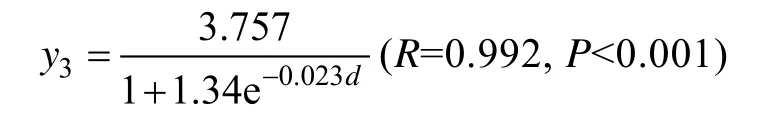
式中y3为厚度,d为生长期。
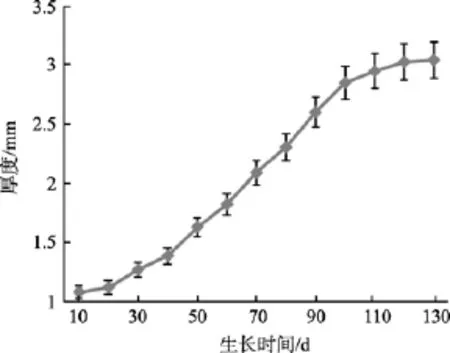
图4 厚度生长曲线Fig. 4 Corelation between the thickness and time
海带在脆嫩期前, 厚度较薄, 相关性较小, 当生长至厚成期, 其厚度增长变快, 夹苗位置与海带厚度的相关性变大(表2), 7月上旬, 海带厚度的相关系数R=0.965时, 得到如下方程:
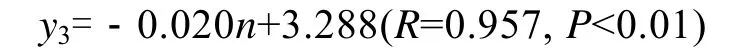
式中y3为厚度,n为夹苗位置。此时, 海带厚度的实测值, 与最佳水层时的期望值的关系为:y′3=0.020(n-1)+x3。
2.4 生长期、夹苗位置对海带鲜质量的影响
海带在厚成期前, 叶片较薄, 其鲜质量变化较慢, 厚成期至成熟期变化最快, 成熟期以后, 速度又放缓, 鲜质量生长曲线(图5)可以表示为:
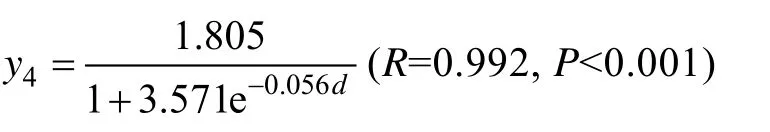
式中y4为鲜质量,d为生长期。
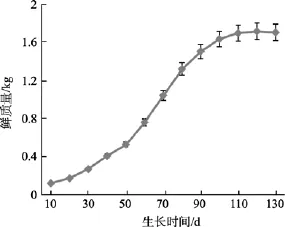
图5 鲜质量生长曲线Fig. 5 Correlation between the fresh weight and time
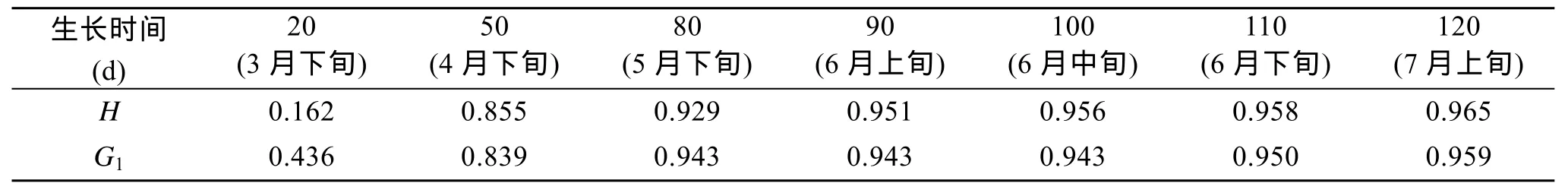
表2 海带的厚度(H)、鲜质量(G1)在不同生长阶段与夹苗位置之间的相关性比较Tab. 2 Correlation between the thickness (H) and fresh weight(G1) of Laminaria japonica at different growth stages and clip seedlings positions
厚成期前, 海带鲜质量较小在水流的作用下多处于悬浮状态, 该阶段与夹苗位置的相关性较低,随着鲜质量的增加, 其相关性逐步提高(表2), 到 7月上旬, 得到夹苗位置与鲜质量的方程:
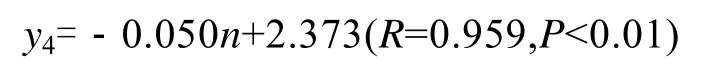
式中y4为鲜质量,n为夹苗位置。此时, 海带鲜质量的实测值, 与最佳水层时的期望值的关系为:y′4=0.050(n-1)+x4。
2.5 夹苗位置对海带干质量的影响
7月上旬, 海带干质量最大, 是收获和海带选种的最佳阶段。对海带的夹苗位置n、长度L(cm)、宽度W(cm)、厚度H(mm)、鲜质量G1(kg)、干质量G2(kg)作出统计(表3), 从统计中可以看出, 宽度、厚度、鲜质量是影响干质量的主要因素。干质量与夹苗位置的方程为:
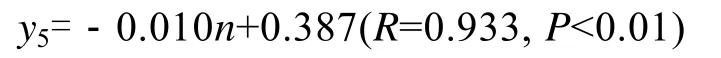
式中y5为干质量,n为夹苗位置。此时, 海带干质量的实测值, 与最佳水层时的期望值的关系为y′5=0.010(n-1)+x5。
3 讨论
3.1 遗传的同源性是准确分析夹苗位置对海带个体生产力影响的关键
在孢子水育苗法中, 精卵随机配对, 即使来自同一株种海带, 海带苗的基因型也存在差异, 如果采用这样的苗种进行本实验, 基因型的差异势必会影响结果。而克隆育苗采用来自同一株雌、雄配子体经过无性繁殖产生的无性繁殖系, 可以保证苗种基因的同一性, 消除了因基因型不同而带来的随机误差。实验中采取了二次夹苗, 是因为前期海带处于凹凸期, 叶片较小、厚度较薄, 海带之间的遮光不明显, 夹苗位置对个体生长力的影响很少; 同时二次夹苗取长、宽相同的海带夹苗可以保证实验材料的同一性, 消除了因实验海带大小不同而带来的误差。这样, 在水文条件相同的同一海域, 夹苗位置就成为环境条件差异的唯一因素。
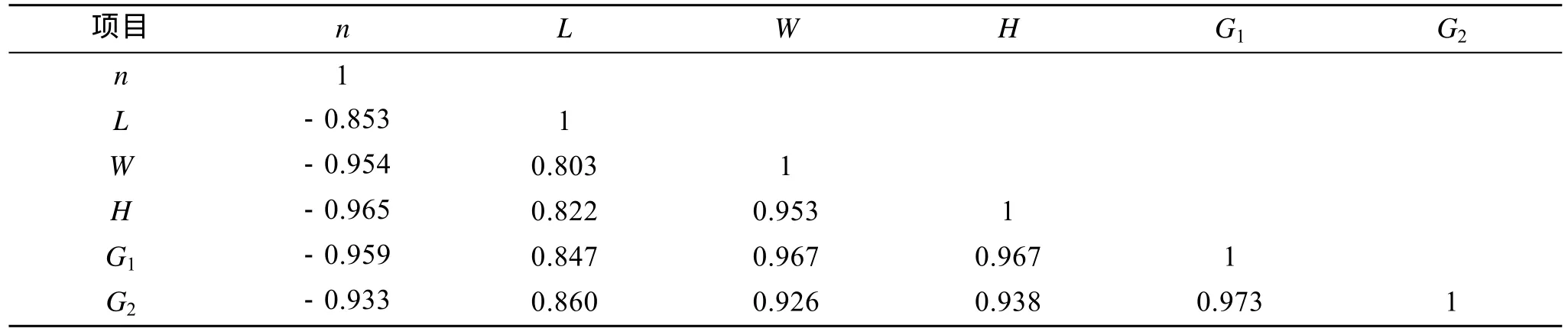
表3 收获后各项指标的相关系数矩阵Tab. 3 Correlation among various indexes after kelp harvest
3.2 夹苗位置对海带个体生产力的影响巨大, 超出品种差异带来的影响
海带育种的目的是为了培育基因型优良的品种,而新品种增产幅度一般不会超过 60%, 如我国早期培育出的“860”和“1170”, 这两个品种能增产 8%~40%[12], 此后中科院又以“860”和“1170”杂交出“海杂一号”, 能够在原基础上增产 6.1%[13], 即使后来采用克隆育苗法育出的高产、耐高温、抗腐烂的“901”海带品种, 其增产幅度也仅仅达到 60%[14]。与夹苗位置比较, 新品种对个体生产力的影响仍然较少,实验结果显示, 在同一苗绳的上水层 4株海带平均干质量(0.36 kg)是下水层 4株海带平均干质量(0.15 kg)的 2倍以上。倘若育苗选种时, 基因型最好的海带位于苗绳下端, 其良好的基因是难以克服环境条件的影响, 表达出超过上水层基因型差的海带的生长性状, 这样很多性状优良的海带会因为水层的限制生产性状表现出来的并不优良, 在选种时丢失,造成种质资源的浪费; 而上水层基因型并不优良的海带由于表现优良可能被选中, 误选对育种工作影响更大。
3.3 水层影响光合作用效率, 进而影响海带鲜干比
海带夹苗位置的不同对海带生产力造成的差异,归根结底是光照的差异[15], 究其原因主要由以下原因引起:第一是水深影响了海带的光合作用。海带随深度的增加, 个体生产力降低; 第二是海带叶片长大以后相互之间的遮光, 也影响其光合作用。Gagn'e等[16]发现, 成体海带叶片组织在 1~15℃范围内达到光合饱和量时, 光照强度约7 500~9 000 lx, 而实验海区的透明度长时间在1m左右, 光强随着水深的变化巨大, 因此水层深浅对光合作用影响较大。同一苗绳水层深的海带, 其光照的不足不仅体现在长、宽等指标上, 鲜干比也随着水层的变深而降低, 由第一株的6.1:1降至最后一株的8.2:1。
3.4 正确选择估算海带个体生产力指标的重要性
选择恰当的指标作为估算海带个体生产力是非常重要的。长度和宽度只能作为衡量海带个体生产力的辅助指标, 在4月中旬之前, 海带植株较小, 在水流的作用下基本上都悬浮于相对一致的水层, 海带长度、宽度与夹苗位置相关性较低, 此时以长宽作为估算海带的个体生产力与夹苗位置的指标误差很大; 6月中旬后, 水温上升(图2), 海带梢部出现不同程度的腐烂, 实验数据显示夹苗位置与长度的相关性由6月上旬0.884降至7月上旬的0.853(表1), 这说明在这一个月中, 海带梢部的腐烂速度逐步大于生长速度。尤其是上水层的海带腐烂现象尤为严重,这样对于 7月份海带选种时以长度为指标估算海带个体生产力显然是不合适的。厚度的增长在凹凸期变化很小, 脆嫩期及厚成期才出现较快的变化, 到了 7月上旬, 海带的厚度与夹苗位置的相关系数达到了 0.965(表2), 但是以厚度作为指标估算海带个体生产力一方面测量难度较大, 另外海带厚度与其他指标相比对光的依赖更大, 上水层光照较好的海带尚可较好的表达出厚度基因, 但下水层光照不足的情况, 海带厚度基因不能得到准确表达。海带栽培的目的就是为了获得海带干品, 因此海带的干质量最能反映不同夹苗位置的海带个体生产力(R=0.933)。虽然实验测得数据, 海带的鲜质量与干质量的相关系数达到了 0.973(表3), 但是由于不同的水层导致的鲜干比出现了一定的差异, 所以采用海带的干质量作为指标能更好的反映出夹苗位置与海带个体生产力的关系:
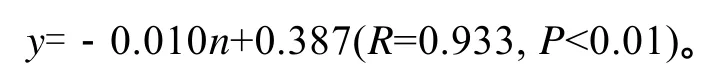
3.5 公式y=0.010(n-1)+x的应用
本实验得出的公式y=0.010(n–1)+x反映了在荣成俚岛湾养殖区, 平养的海带不同夹苗位置在 7月上旬实测干质量与在最上层位置养殖时的期望值之间的关系, 这个公式既反映了夹苗位置对海带个体生产力的影响, 同时又可以应用于海带育种中的选种工作中:对于实验中表现较好的个体进行测量,并记录其位置, 然后代入公式计算出每个样本的期望值, 由于该期望值消除了环境因素带来的差异,把不同夹苗位置上的海带的干质量统一为最佳位置的干质量期望值, 可以进行量化比较, 这样就避免了选种中的漏选和误选, 极大提到选种工作的准确度和效率。
4 小结
干质量最能反映夹苗位置与个体生产力的相关关系, 夹苗位置对个体生产力影响可以用公式y=–0.010n+0.387(R=0.933,P<0.01,y代表海带个体生产力)来表达; 不同夹苗位置海带个体生产力的期望值可以用下面方程表示:y=0.010(n-1)+x(其中y为期望值,x为实测值), 该公式表达了不同基因型的海带采取浮筏式平养时, 在排除夹苗位置因素干扰后其基因型应该表达出的个体干质量期望值。
[1]曾呈奎, 王素娟, 刘思俭, 等.海藻栽培学[M].上海:上海科学技术出版社, 1985.
[2]Valrie A. Gerard.Environmental stress during early development of kelp sporophytes(Laminaria saccharina)[J]. how long do effects persist?1997, 9:5-9.
[3]Gerard V A. Ecotypic differentiation in light-related traits of the kelpLaminaria saccharina[J]. Mar Biol,1988, 97:25–36.
[4]Gerard V A. Ecotypic differentiation in the kelpLaminaria saccharina:phase-specific adaptation in a com-plex life cycle[J]. Mar Biol, 1990, 107:519–528.
[5]Zhang Q S, Qu S C, Cong Y Z, et al. High throughput culture andgametogenesis induction ofLaminariajaponicagametophyte clones[J]. Journal of Applied Phycology, 2008, 20:205-211.
[6]张定民, 廖国荣. 沿岸流与海带养殖关系的研究[J].山东海洋学院学报, 1986, 16(4):180-185.
[7]孙国玉.50 年代我国海带栽培研究的几件事[J].海洋科学, 1993, 2:69-70.
[8]王素娟.海藻生物技术[M].上海:上海科学技术出版社, 1994:104-111.
[9]李志凌, 张全胜, 杨迎霞, 等.海带配子体克隆大规模培养技术的研究[J].齐鲁渔业, 2003, 20(5):1-3.
[10]李凤晨, 李豫红. 海带筏式养殖技术要点[J].河北渔业, 2003, 3:17-20.
[11]Dokulil M. Optical properties, colour and turbidity[C]//Loffler H. Neusiedlersee:The limnology of a shallow lake in cenrrtral Europe. The Hague:Dr W Junk bv Publishers, 1979:160-161.
[12]青岛海洋水产研究所藻类养殖组, 中国科学院海洋研究所海藻遗传育种组.高产高碘海带新品种的培育[J].遗传学报, 1976, 3(3):224-230.
[13]缪国荣, 丁夕春.我国海带育种的成果及其评价[J].齐鲁渔业, 1985, 2:15-18.
[14]张全胜, 刘升平, 曲善村, 等.“901”海带新品种培育的研究[J].海洋湖沼通报, 2001, 2:46-53.
[15]Aguilera J, Gordillo F J L, Karsten U, et al. Light quality effect on photosynthesis and efficiency of carbon assimilation in the red algaPorphyra leucosticta[J].Journal of Plant Physiology, 2000, 157:86-92.
[16]Gagne J A, Mann K H.Comparison of growth strategy inLaminariapopulations living under differing seasonal patterns of nutrient availability[J].Proc Int Seaweed Symp, 198110; 297-302.
The correlation analysis of individual productivity and clip seedlings position ofLaminaria japonicaon raft culture
REN Wei, YUAN Zhu-tao, LIU Sheng-ping
(Qingdao Agricultural University, Qingdao 266109,China)
Feb.,23,2012
Laminaria japonica; raft culture; individual productivity; seedling clamping position; seeds selection
For the raft culture breedingLaminaria japonica, individual productivity was closely related to clip seedlings position.L. japonicain upper water grew better. Using the method of collecting germplasm according to character quality, the germplasm we needed accounted for only about 10% in all ofL. japonica.The experiment materials were cloned seedlings from the same gametophyte clones, and made correlation analysis between the individual productivity and clip seedlings position. Comparison among length, width, thickness, fresh weight and dry weight showed that the dry weight could reflect individual productivity preferably. We derived an equation,y=-0.010x+0.387(R=0.933,P<0.01). This equation expressed the clip seedlings position influence individual productivity. In selecting seeds ofLaminariabreeding, we could calculate the expectation values from the experimental values measured at the same clip seedlings position by the equation,y=0.010(n-1)+x, enabling the data in different seedlings position to be comparable, eliminating the environment factors that influence the individual productivity ofL. japonicaand avoiding missing or mistakenly selection of seeds.
Q949 文献标识码:A 文章编号:1000-3096(2012)09-0122-06
2012-02-23;
2012-08-22
农业部948资助项目(Z-1025)
任伟(1986-), 男, 硕士, 主要从事海藻生物学方面研究,E-mail:renwei-post@163.com; 刘升平, 通信作者, 男, 教授, 硕士生导师, E-mail:lshengping88@yahoo.com.cn
梁德海)
