渤黄海营养盐结构及其潜在限制作用的时空分布
2012-01-12谢琳萍王保栋
谢琳萍, 孙 霞, 王保栋, 辛 明
(1. 国家海洋局 第一海洋研究所 海洋生态研究中心, 山东 青岛 266061; 2. 海洋生态环境科学与工程国家海洋局重点实验室, 山东 青岛266061)
渤黄海营养盐结构及其潜在限制作用的时空分布
谢琳萍1,2, 孙 霞1,2, 王保栋1,2, 辛 明1,2
(1. 国家海洋局 第一海洋研究所 海洋生态研究中心, 山东 青岛 266061; 2. 海洋生态环境科学与工程国家海洋局重点实验室, 山东 青岛266061)
根据2006-2007年4个季节的现场调查资料, 分析探讨了渤海和黄海营养盐结构分布变化特征及其对浮游植物生长的潜在的限制状况。结果表明, 渤黄海水体Si/N/P比值均偏离Redfield比值, 季节变化明显; 春夏冬季N/P和Si/P比值由近岸向远岸海域递减, 高值区主要分布在黄河口、鸭绿江口及苏北近岸, 秋季上层水体N/P和Si/P比值的分布趋势有所不同, 高值区主要分布在南黄海的中部海域。受陆源输入的影响, 近岸特别是河口区N/P和Si/P比值均较高, 温跃层的生消变化和生物活动调控着黄海中部海域营养盐结构的变化。渤黄海浮游植物生长主要受P的潜在限制, 部分季节受N、Si的潜在限制; 营养盐限制状况存在着明显的时空变化及不同营养盐的同时或交替限制的现象。
营养盐结构; 营养盐限制; 渤海; 黄海
营养盐是浮游植物生命活动的物质基础, 浮游植物按一定比例(Redfield比值)摄取营养物质维持自身的物质和能量代谢[1], 适宜的N/P、Si/N和Si/P比值有利于浮游植物的生长和繁殖, 反之则会造成营养结构失调, 浮游生物的种群结构改变, 甚至引发赤潮[2-3]。低N/P比值或较高浓度的磷酸盐有利于硅藻的生长, 而高 N/P比值或较高浓度的氮盐则有利于甲藻的生长; 硅酸盐对硅藻的生长非常重要, 低N/Si比值或较高浓度的硅酸盐可使硅藻在与甲藻竞争中占据优势[4]。一般来说, 沿岸和较封闭海域易发生P限制, 如:地中海[5-6]、芬兰西南群岛附近海域[7];在水交换较好的外海和大洋, 多发生 N限制, 如波罗地海[8]和Kaneohe[9]湾; 在咸淡水交界的河口地带,易出现营养盐的同时或交替限制[10]。营养盐的限制具有时间变化, 在同一海区往往会出现N、P营养盐限制的季节性交替变化[11]。近年来Si限制的报道有所增多, Fe、Mn等微量元素的缺乏亦可限制浮游植物的生长[12-13]。
渤海、黄海是我国重要的半封闭陆架海区, 具有陆源输入量大、海水交换率小、初级生产力高等特点。随着沿岸经济社会的发展, 工农业和城市生活污水的排放, 使海区营养盐浓度和结构发生了明显的变化[2,14-15], 而营养盐结构的变化不仅反映了营养盐状况的变化, 同时反映了海水中营养盐的再生、循环机制的改变, 对海区生态环境产生较大的影响[2-3,15-17]。本文根据2006-2007年对黄渤海4个季节的现场调查资料, 对整个海区不同季节营养盐结构的分布、变化及其对浮游植物生长的潜在限制状况进行分析, 探讨渤黄海营养盐结构现状及其对浮游植物生长的影响。
1 调查与方法
本文所用资料来源于908成果集成项目“中国近海化学调查研究”, 其中渤海数据是由国家海洋局北海分局提供; 北黄海数据参考文献报道[15]。
本文所用数据的调查时间分别于2007年4月(春季)、2006 年 7 月(夏季)、2007 年 10 月(秋季)和 2007年1月(冬季), 每个航次345个调查站位, 调查站位分布见图1所示。
现场用Seabird CTD对温度、盐度、深度等参数进行测定, 水样用 Niskin采水器按照 0、10、30 m和底层采集, 水样采集后经 0.45μmol/L 醋酸纤维滤膜过滤后, 用于营养盐分析。其中无机磷(PO4-P)用原磷钼蓝法, 活性硅酸盐(SiO3-Si)用硅钼蓝法, 亚硝酸盐(NO2-N)用盐酸萘乙二胺法, 硝酸盐(NO3-N)用 Cd-Cu还原法, 铵盐(NH4-N)用次溴酸盐氧化法。无机氮(DIN)= NO3-N+NO2-N+NH4-N。
2 结果与讨论
2.1 渤黄海营养盐比值的季节变化
从表1可以看出春季N/P和Si/P比值的变化幅度较大, 最高值分别为786.6和399.2。Si/N比值数据离散性不大, 均值为1.23; 夏季N/P、Si/N和Si/P比值均高于春季; 秋季N/P比值均值最大, 达四季之最, 而Si/N和Si/P比值则比夏季小; 冬季N/P、Si/N和Si/P比值不大, 变化幅度也很小。综上所述, N/P比值季节变化为秋季>夏季>春季>冬季; Si/N比值的季节变化为夏季>冬季>秋季>春季; Si/P比值的季节变化为夏季>秋季>春季>冬季。
对上层水体营养盐结构进行进一步分析, 由图2可见春季大部分站位 Si/N比值小于 Redfield比值,而Si/P比值, N/P比值则大于Redfield比值。夏季上层水体约有23%的站位N/P比值小于Redfield比值,大部分站位的Si/N和Si/P比值大于Redfield比值(分别为78%和86%)。秋季仅有极小数站位的N/P、Si/P比值小于Redfield比值(分别为8.9%和4%), 约有1/3站位的Si/N比值小于Redfield比值。冬季N/P、Si/N和 Si/P比值的数据离散性不大, 大部分站位 N/P/Si比值高于Redfield比值。

表1 渤黄海营养盐结构的统计结果Tab. 1 Statistic results of the nutrient structure in the Baohai Sea and the Yellow Sea
2.2 渤黄海营养盐结构的平面分布
春季, 上层水体 N/P比值的高值区主要出现黄河口、鸭绿江口及苏北近岸, 低值区则出现在渤黄海中部海域, 整体呈现由近岸向远岸递减的变化趋势(图3a)。其中黄河口和鸭绿江口 N/P比值很高(100~779.5), 苏北沿岸 N/P比值也在 25以上, 而黄海中部海域N/P比值含量较低(<16)。底层N/P比值的分布趋势与上层水体类似, 最高值仍出现在莱州湾附近海域, 达786.6, 鸭绿江口N/P比值仍然较高,但高值范围比上层明显减小, 山东半岛附近海域N/P比值相对较低(<10)。Si/N比值在上层水体的分布为黄海中部海区较高(>2), 渤海和黄海近岸 Si/N比值均较低(<1)。底层 Si/N比值的分布趋势与上层水体基本一致, 不再陈述。水体 Si/P比值的变化范围较大(1.20~399.2), 山东半岛附近海域 Si/P比值较低, 黄河口附近海域较高, 其他海域 Si/P比值均大于10。
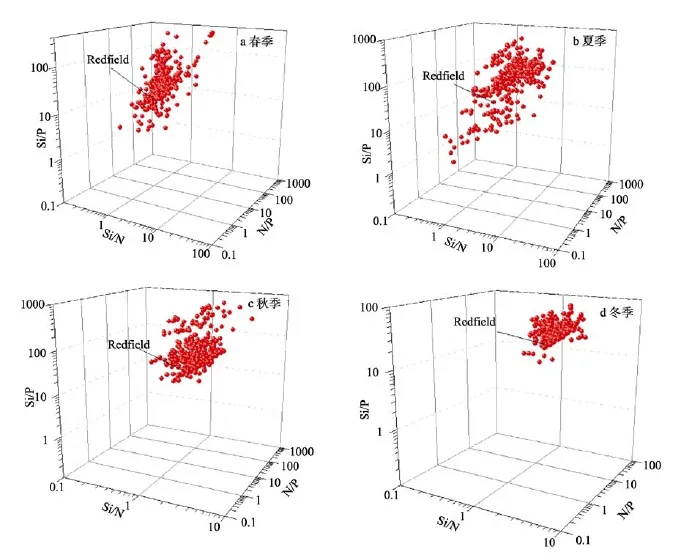
图2 上层水体营养盐结构的三维散点图Fig. 2 Three-dimensional scatter plots of the nutrient structure in the upper water
入春以后, 随着气温的升高, 江水解冻后将高DIN、低PO4-P的淡水排放入海[18-19], 造成黄河口、鸭绿江口等河口区域N/P和Si/P比值较高。青岛近岸海域受外海冷水的影响[20], 水体温度较低(8℃),生物活动能力相对较弱, 对水体营养盐的消耗相对较小, 海区N/P和Si/P比值较低。此外, 初春叶绿素a高值基本分布在温度高的近岸海区[21], 黄海中部海域叶绿素 a的含量较低, 生物活动对营养盐的消耗相对较小, N/P比值较低。
夏季, 近岸上层水体高 N/P比值的特性更加明显, 高值区范围较春季有所扩大, 呈现由近岸向外海递减的变化趋势, 其中莱州湾及海州湾南部沿岸N/P比值均在200以上, 长江口北部海域N/P比值也较高(>100), 南黄海的中北部海域 N/P比值较低(<12)(图3b)。上层水体Si/N比值高值区主要出现在渤海的中部和青岛附近海域, 北黄海深水区和南黄海大部分海区 Si/N比值较低(<2)。Si/P比值的分布趋势与N/P极为相似, 高值区主要出现在渤海近岸、北黄海近岸、浙江沿岸和长江口北部海域。底层N/P、Si/N和 Si/P比值的分布趋势与上层水体基本一致,不再赘述。
综合来看, 受高N/P、高Si/P含量的黄河冲淡水影响, 黄河口附近海域呈现高N/P、高Si/P比值的特点, 而受陆源输入较少的南黄海中部海域则呈现低N/P、低Si/N和低Si/P比值的特点。黄海南部高N/P、低 Si/N含量(N/P>100, Si/N<1)的长江冲淡水在偏南季风和台湾暖流的影响下, 在 122°30′附近左转向北或东北进入南黄海西南部海域[22-23], 受其影响长江口附近海域N/P和Si/P比值较高。受射阳河输入和苏北沿岸流的影响, 海州湾以南(34°N)近岸海域N/P和Si/P比值也比较高。
秋季, 高 N/P比值含量的水体占据着南黄海中部的大部分海区, 并呈舌状由东部外海向近岸扩展,高值中心达500以上。此外, 黄河口、海州湾附近海域也出现了小范围的N/P高值(图3c)。底层水体N/P的分布趋势与上层水体有一定的差异, 高值区主要出现在近岸, 黄海中部海域N/P比值较低, 该部分正好与上层水体的 N/P比值高值区相反。渤黄海上层水体中 Si/N比值不高(1~5), 近岸较低, 中部海域稍高(图4a), 底层 Si/N比值的分布趋势与上层基本一致, 不再赘述。水体中 Si/P比值的分布趋势与 N/P比值相似, 不再赘述(图4c)。
综合来看, 秋季黄河口附近海域仍呈现高N/P、低 Si/N比值的特点, 南黄海中部上层水体则呈现高N/P、高Si/P比值的特点, 而底层则表现为低N/P、Si/P比值, 该区域与夏季黄海冷水团所在位置基本一致, 说明随着温跃层的减弱, 水体上下层混合加强, 黄海冷水团内丰富的营养盐对上层水体起到一定的补充作用, 适宜的温度和光照条件, 促进了上层水体浮游植物的生长, 加大了对水体营养盐特别是PO4-P的消耗, 使上层水体N/P、Si/P比值较高, 而底层水体浮游植物活动较弱, 水体N/P、Si/P比值较低。

图3 上层水体N/P比值的平面分布Fig. 3 The horizontal distributions of N/P ratios in the upper water

图4 秋季和冬季上层水体Si/N、Si/P比值的平面分布Fig. 4 The horizontal distributions of Si/N and Si/P ratios in the upper water in autumn and winter
冬季:水体垂直混合加强, 上下层水体性质趋于一致, 水体营养盐结构分布变化不大。N/P比值:高值区主要分布在黄河口近岸、海州湾及苏北沿岸,低值区则主要出现在渤海、北黄海中部及南黄海南部海域(图3d)。Si/N比值高值区主要出现在渤海的中部、青岛近岸海域, 低值区(小于 1)则主要出现在苏北沿岸(图4b)。Si/P比值在黄河口附近海域和苏北沿岸水体稍高、北黄海的东部稍低, 整个海域 Si/P比值分布差异不明显(图4 d)。
2.3 营养盐结构对浮游植物生长的限制
2.3.1 浮游植物生长的营养盐限制标准
国内外众多学者对浮游植物生长的营养盐限制标准进行了研究[24-27](表2), Nelson等[24]利用浮游植物生长动力学得出浮游植物所能利用营养盐的最低浓度, 即营养盐的绝对限制标准; Justic等[25]根据Si/N/P比值提出营养盐的相对限制标准; Dortch and Whitledge[27]结合营养盐的绝对和相对限制标准提出了一套营养盐的限制标准, 但这些标准或不够全面或部分参数范围太宽, 为了能更好地反映浮游植物生长的营养盐限制情况, 本文综合了上述各种判别标准, 改进了 Dortch等[27]提出的营养盐限制标准,将N和Si的限制标准进行了部分调整, 具体如表2所示。

表2 国内外营养盐限制标准的主要研究结果Tab. 2 The nutrient limitation standard
2.3.2 黄渤海浮游植物生长的营养盐限制状况
根据上述营养盐限制标准得出调查海域营养盐的限制情况(表3, 图5), 春季 N潜在限制站位以南黄海最多, 主要分布在山东半岛的南部海域, 而P潜在限制站位则多分布在北黄海, 受限站位占北黄海调查站位的12.1%; Si潜在限制站位主要集中在南黄海的海州湾附近海域(14个站位), 渤海和北黄海浮游植物生长基本不受Si的限制。
夏季调查海区浮游植物生长以P潜在限制为主,仅部分海域出现小范围的 N和 Si的潜在限制。具体而言, 南黄海 N潜在受限站位最多(17个站位),分布特性由春季山东半岛南部海域的集中分布转为南黄海中部海域的零星分布; P潜在限制状况较春季更为严重, 在黄河口、北黄海和海州湾南部海域均出现较大范围的P潜在限制, 以北黄海最为突出, 有 58%的调查站位受到 P的潜在限制; 南黄海中部海域出现部分站位Si潜在限制, 渤海和北黄海无Si潜在限制的现象。秋季水体营养盐状况有所好转, 浮游植物生长基本不受 N、Si潜在的限制, 主要受 P潜在的限制, 受限站位主要出现在南黄海,占南黄海调查站位的35%。从分布趋势上来看, P潜在限制区域与夏季相比发生了明显的变化, 限制区域主要分布在黄海中部和海州湾附近海域。冬季水体营养盐状况较好, 浮游植物生长基本不受营养盐的限制, 仅在海州湾附近海域出现零星站位N或P的潜在限制。
从海区上来看:渤海营养盐含量丰富, 仅黄河口附近海域在夏季受P的潜在限制。
北黄海在春夏秋季节浮游植物生长均受到 P的潜在限制, 夏季尤为突出。春季高N/P比值的陆源输入造成北黄海水体 N/P比值较高[19], 而浮游植物的生物活动加大了对营养盐的消耗使水体 PO4-P含量较低(<0.1μmol/L), 造成水体浮游植物生长受到 P的潜在限制。夏季调查海区仍存在着高 N/P比值的陆源输入和浮游植物对营养盐的消耗, 但由于温跃层的形成阻碍了底层水体营养盐对上层水体的补充,导致北黄海浮游植物生长受 P潜在限制的区域面积较春季大。秋季随着温跃层的减弱, 富含营养盐的下层水体的上涌, 在一定程度上缓减了上层水体缺 P的状况, 海区浮游植物生长受 P潜在限制的区域面积较夏季有所减少, 受限制的区域仅出现在北黄海的深水区。
南黄海浮游植物生长主要受 P的潜在限制, 以秋季最为突出, 春夏季节小部分海域浮游植物生长受N或Si的单独和同时潜在限制。秋季随着温跃层的减弱, 水体上下层混合加强, 冷水团内的丰富的营养盐对上层水体起到一定的补充, 但南黄海中部大部分海域 N/P和 Si/P比值仍然很高(N/P>100,Si/P>200), 浮游植物生长受 P的潜在限制。从上面论述可以看出营养盐对浮游植物生长的潜在限制作用具有季节变化性, 常发生几种营养盐同时或交替限制的现象, 与国内外许多学者的研究结果相一致[11,15]。

表3 上层水体营养盐限制站位的统计结果Tab. 3 Statistical results of nutrient limitation stations in the upper water

图5 上层水体营养盐限制的平面分布Fig. 5 The distributions of nutrient limitation stations in the upper water
3 结论
(1)渤黄海水体中N/P、Si/N和Si/P比值均偏离Redfield比值, 季节变化显著, N/P比值的季节变化为:秋季>夏季>春季>冬季; Si/N比值的季节变化为夏季最高, 冬季次之, 春秋季较小; Si/P比值的季节变化为夏季>秋季>春季>冬季。
(2)春夏冬季N/P和Si/P比值由近岸向远岸海域递减, 高值区主要分布在黄河口、鸭绿江口及苏北近岸, 南黄海中部海域 N/P 和 Si/P比值较低, 秋季上层水体 N/P和 Si/P的分布与其他季节不同, 高值区主要出现在南黄海的中部和莱州湾附近海域。
(3)高N/P比值的陆源输入是造成近岸特别是河口区高N/P、Si/P的主要原因, 而生物活动和温跃层的生消变化是影响黄海中部营养盐结构季节变化的主要因素。
(4)渤黄海浮游植物生长主要受 P的潜在限制,部分季节受N和Si的潜在限制。营养盐限制的区域存在明显的季节变化, 存在着不同营养盐同时或交替限制的现象。
[1]Redfield A C. The biological control of chemical factors in the environment [J].American Scientist,1958,46:205-222.
[2]高生泉, 林以安, 金明明, 等.春、秋季东、黄海营养盐的分布变化特征及营养结构[J].东海海洋, 2004,22 (4):38-50.
[3]蒲新明, 吴玉霖, 张永山. 长江口区浮游植物营养限制因子的研究Ⅱ.春季的营养限制情况[J]. 海洋学报,2001, 23(3):57-65.
[4]李京. 东海赤潮高发区营养盐结构及对浮游植物优势种演替的作用研究[D]. 中国海洋大学, 2008, 5-7.
[5]Zohary T, Robarts R D. Experimental study of microbial P limitation in the eastern[J]. Limmology and Oceanography, 1998, 43(3):387-395.
[6]Thingstad T F, Zweifel U L, Rassoulzadegan F.P limitation of heterotrophic bactena and phytoplankton in the northwest Mediterranean[J]. Limmology and Oceangraphy, 1998, 43(1):88-94.
[7]Kirkkala T, Helminen H, Erkkilä A. Variability of nutrient limitation in the Archipelago Sea, SW Finland[J].Hydrobiologia, 1997, 363(1-3):117-126.
[8]Maestnm S Y, Balode M, Béchemin C, et al. Nitrogen as the nutrient limiting the algal growth potential for summer natural assemblages in the Gulf of Riga,Eastem Balitic Sea[J].Plankton Biology and Ecology,1999, 46(1):1-7.
[9]Lamed S T. Nitrogen versus phosphorus-limited growth and source of nutrients for coral reef macroalgae[J].Marine Biology, 1998, 132(3):409-421.
[10]王保栋. 黄海和东海营养盐分布及其对浮游植物的限制[J]. 应用生态学报, 2003, 14(7):1122-1126.
[11]Fisher T R, Peele E R, Ammerman J W, et al. Nutrient limitation of phytoplankton in Chesapeake Bay[J].Mar.Ecol. Prog. Ser, 1992, 82:51-63.
[12]Dugdale R C, Wilkerson F P, Minas H J. The role of a silicate pump in driving new production[J].Deep-Sea Res, 1995, 42:697-719.
[13]Martin J H. Iron as a limiting factor in oceanic productivity. In:Falkowski PG & Woodhead AD eds. Primary Productivity and Biogeochemical Cycles in the Sea[M].New York:Plenum Press, 1992, 123-137.
[14]蒋红、崔毅、陈碧鹃, 等. 渤海近20年来营养盐变化趋势研究[J]. 海洋水产研究, 2005, 26(6):61-67.
[15]张辉, 石晓勇, 张传松, 等. 北黄海营养盐结构及限制作用时空分布特征分析[J]. 中国海洋大学学报,2009, 39(4):773-780.
[16]王修林, 孙霞, 韩秀荣, 等. 2002年春、夏季东海高发区营养盐结构及分布特征的比较[J]. 海洋与湖沼,2004, 35(4):323-330.
[17]韦桂峰. 广东大亚湾西南部海域营养盐结构的长期变化[J]. 生态科学, 2005, 24(1):1-5.
[18]张晓晓, 姚庆祯, 陈洪涛, 等. 黄河下游营养盐浓度季节变化及其入海通量研究[J].中国海洋大学学报,2010, 40(7):82-88.
[19]熊辉, 于志刚, 陈洪涛, 等. 鸭绿江口溶解态 P、Si的地球化学研究[J]. 海洋环境科学, 1999, 18(2):1-6.
[20]邹娥梅, 郭炳火, 汤毓祥. 1996年春季南黄海水文特征和水团分析[J]. 海洋学报, 2000, 22(1)17-26.
[21]胡好国, 万振文, 袁业立.南黄海浮游植物季节性变化的数值模拟与影响因子分析[J], 海洋学报, 2004,26(6):74-88.
[22]李玲玲, 于志刚, 姚庆祯, 等. 长江口海域营养盐的形态和分布特征[J]. 水生态学杂志, 2009, 2(2):15-20.
[23]邹娥梅, 郭炳火, 汤毓祥, 等. 南黄海及东海北部夏季若干水文特征和环流的分析[J]. 海洋与湖沼, 2001,32(3):341-348.
[24]Nelson D M, Brzezinski M A. Kinetics of silicic acid uptake by natural diatom assemblages in two Gulf Stream warm-core rings[J]. Marine Ecology Progress Series, 1990, 62:283-292.
[25]Justic D, Rabalais N N, Turner R E. Stoichiometry nutrient balance and origin of coastal eutrophication [J].Marine Pollution Bulletin, 1995, 30:41-46.
[26]胡明辉, 杨逸萍, 徐春林, 等.长江口浮游植物生长的磷酸盐限制[J]. 海洋学报, 1989, 11(4):439-443.
[27]Dortch Q, Whitledge T E. Does nitrogen or silicon limit phytoplankton production in the Mississipi River plume and nearby regions[J]. Continental Shelf Research,1992, 12:1293-1309.
Temporal and spatial distributions of nutrient structure and potential nutrient limitation in the Bohai Sea and the Yellow Sea
XIE Lin-ping1,2, SUN Xia1,2, WANG Bao-dong1,2, XIN Ming1,2
(1. Marine Ecology Research Centre, the First Institute of Oceanograghy, SOA, Qingdao 266061, China; 2. Key Lab of Science and Engineering for Marine Ecological Environment, SOA, Qingdao 266061, China)
Jan.,13,2012
nutrient structure; nutrient limitation; Bohai Sea; Yellow Sea
Based on the seasonal field observation during 2006 -2007, distributions of nutrient structure and their potential limitation on phytoplankton growth in the Bohai Sea and the Yellow Sea were discussed. The results showed that ratios of N/P, Si/N and Si/P varied greatly in different seasons, and deviated from Redfield ratios. The ratios of N/P and Si/P decreased from inshore to offshore area in spring, summer and winter, while showed the different pattern in the upper water in autumn, higher values of N/P and Si/P ratios were observed in the middle of the southern Yellow Sea. Due to the influence of terrestrial runoff, the inshore waters especially at the estuary area presented very high N/P and Si/P ratios. Changes of thermocline and biological activities controlled the nutrient structure in the central area of the Yellow Sea. The primary production might be limited by phosphate in a considerable portion of the Bohai Sea and the Yellow Sea. The potential nutrient limitation showed temporal and spatial variations, and the phytoplankton were usually limited by single or multiple nutrients at the same time or subsequently.
P374 文献标识码:A 文章编号:1000-3096(2012)09-0045-09
2012-01-13;
2012-04-16
中国近海化学调查研究项目(908-JC-1-03)
谢琳萍(1982-), 女, 山东烟台人, 硕士, 研究实习员, 主要从事海洋生源要素的生物地球化学研究, 电话:0532-88977135, Email:xlp@fio.org.cn; 王保栋, 通信作者, 研究员, 电话:0532-88962016,Email:wangbd@fio.org.cn
致谢:感谢国家海洋局北海分局、中国海洋大学和国家海洋局第一海洋研究所的所有参与现场调查和数据分析工作的老师和同学。
康亦兼)
