紫扇贝和海湾扇贝杂交家系的生长和通径分析
2012-01-11李建立王春德李朝霞蔡德泉张金盛
李建立, 王春德, 李朝霞, 蔡德泉, 张金盛
(1. 青岛农业大学, 山东青岛266109; 2. 威海市海洋与渔业局, 山东威海264200; 3. 招远市夏甸农业技术推广站, 山东烟台 265415)
紫扇贝和海湾扇贝杂交家系的生长和通径分析
李建立1, 王春德1, 李朝霞1, 蔡德泉2, 张金盛3
(1. 青岛农业大学, 山东青岛266109; 2. 威海市海洋与渔业局, 山东威海264200; 3. 招远市夏甸农业技术推广站, 山东烟台 265415)
以紫扇贝(Argopecten purpuratus)和海湾扇贝(Argopecten irradians irradians)为亲本, 建立了紫扇贝自交(ZZ)、海湾扇贝自交(HH)、紫扇贝(卵)×海湾扇贝(精)(ZH)和海湾扇贝(卵)×紫扇贝(精)(HZ)共 4个试验家系。在养殖 60、127 d时测定壳高、壳长、壳厚和体质量, 并分析各养殖时期影响体质量的主要因素。结果表明: 在各养殖时期, 2杂交家系子代的壳高、壳厚以及体质量均表现出生长优势(P<0.01), 因此认为通过紫扇贝和海湾扇贝杂交方式是改善扇贝的良好方法。通径分析的结果表明, 对于自交家系, 壳长是影响体质量的主要影响因素, 但对于杂交家系HZ来说, 壳长(60 d)和壳厚(127 d)对体质量的决定作用最大; 在杂交家系ZH, 60 d时壳长和壳厚的协同作用对体质量的决定作用最大, 127 d时壳高的决定作用最大。因此对于两个杂交家系的选育, 应按照不同性状对体质量的决定系数选择各时期的优势性状, 以使子代的杂种优势充分表现。
紫扇贝(Argopecten purpuratus); 海湾扇贝(Argopecten irradians irradians); 杂交家系; 杂交优势; 通径分析
海湾扇贝(Argopecten irradians irradians)自1982年引入中国以来深受养殖业户的欢迎, 并形成了中国水产养殖业一个新的发展高潮[1]。但是, 近年来随着养殖领域的逐渐扩大和养殖历史的增长, 病害频发、个体小型化等问题凸现, 导致商品贝的规格越来越小。另外, 种质退化也导致育苗养殖过程中较高的死亡率[2-3]。尽管从引种复壮等方面做了大量的工作,同时取得了一定的成效[4-8]。但是仍需要从种质入手,借助于遗传改良及育种技术, 培养出一种抗逆性强、生长快、适应性强的扇贝新品种。
紫扇贝(Argopecten purpuratus) 是原产于南太平洋的一种速生型中型扇贝[9]。紫扇贝的壳较宽, 闭壳肌肥大, 出肉率高, 且味道鲜美, 壳形优美, 因此紫扇贝商品价值高。紫扇贝于2008年成功地从秘鲁引进, 并成功地在家系水平进行了紫扇贝与海湾扇贝的种间杂交实验[10-11]。体质量是扇贝的主要经济性状之一, 是选择育种的一个重要指标, 同时也是所有经济性状的综合体现。因此, 找出影响体质量的主要目标性状, 明确选育目的, 具有重要的现实意义。近年来, 多元回归和通径分析已在水产动物鱼类、虾类和贝类研究中得到广泛应用, 研究内容主要集中在生长性状对体质量的影响[12-18]。为了获得通过遗传改良的新品系, 2009年, 采用同样的方法建立了1个紫扇贝(卵)×海湾扇贝(精)杂交家系和1个海湾扇贝(卵)×紫扇贝(精)杂交家系。通过比较2个杂交家系与紫扇贝和海湾扇贝自交家系之间经济性状的差异, 并通过通径分析方法, 确定家系中各形态性状对体质量的相对重要性, 以满足生产的需要。
1 材料与方法
1.1 实验材料
2009年 5月, 采用紫扇贝与海湾扇贝种间杂交的方法[10], 获得两个杂交家系和两个自交家系。
ZH: 紫扇贝(卵)×海湾扇贝(精);
HZ: 海湾扇贝(卵)×紫扇贝(精);
ZZ: 紫扇贝(卵)×紫扇贝(精);
HH: 海湾扇贝(卵)×海湾扇贝(精)。
亲本的规格见表 1。分别隔离培养 4个家系,幼虫培育过程中控制相同密度; 幼虫附着后, 及时分苗并控制各组密度; 2009年7月, 各组扇贝的养成过程均在青岛小口子自然海区扇贝笼中进行,每层30只(每笼10层), 养成实验至 2009年 11月底结束。

表1 实验亲本的大小和体质量Tab.1 The morphological measurements of brood stocks (n=50)
1.2 实验方法
在养成阶段的第60天和第127天, 分别从各实验家系中随机抽取50只扇贝, 用游标卡尺测量每只扇贝的壳高、壳长和壳厚, 电子天平秤量扇贝的体质量。
1.3 分析方法
相关数据分析采用 SPSS 13.0 统计分析软件处理。对于数据比较结果, 当P<0.05 时为差异显著,P<0.01时为差异极显著。用下面的公式来计算杂种优势(Heterosis):

公式中的 VF1代表杂交子一代(ZH或 HZ)性状的平均值,VZZ和VHH分别代表ZZ和HH性状的平均值。计算各性状平均数和标准差, 获得各性状表型参数统计量, 对各生长参数进行方差分析、显著性检验和Duncan 多重比较分析、各性状间表型相关分析和形态性状各指标对体质量的通径分析。在表型相关分析的基础上, 根据通径系数原理得到各形态性状对体质量的通径系数, 分析各性状分别对体质量的直接和间接影响, 并求得决定系数[12]。
2 结果
2.1 子代的生长性状比较
各组在60 d 时生长性状的生长情况见表2, 两个杂交家系(HZ和ZH)之间的壳高、壳长、壳厚和体质量差异均不显著(P>0.05), 而ZH和HZ的壳高、壳长、壳厚和体质量极显著高于ZZ(P<0.01), HH的体质量和ZH与HZ相比差异不显著(P>0.05), HH的壳高、壳长、壳厚和体质量极显著高于ZZ(P<0.01)。各组在127 d时生长性状的生长情况见表3, ZH的壳高、壳长、壳厚和体质量和 HZ相比差异不显著(P>0.05), ZH和HZ的壳高、壳长、壳厚和体质量极显著高于ZZ和HH(P<0.01), HH的壳高、壳长、壳厚和体质量极显著高于ZZ(P<0.01)。
2个杂交家系的杂种优势率见表3, 随着养成时间的增加, ZH和HZ各生长性状的杂种优势率有增大的趋势。在60 d和127 d , ZH和HZ的杂种优势率大小依次是体质量>壳长>壳高>壳宽。在 60 d时, ZH的体质量杂种优势率最大, 是32.37%, HZ的体质量杂种优势率是27.75%, ZH的体质量和壳高的杂种优势率高于HZ。在127 d 时, ZH的体质量杂种优势率最大, 是77.95%, 比HZ的高, ZH壳高、壳长和壳宽的杂种优势率低于HZ。

表2 各子代家系的生长指标Tab. 2 The growth parameters of the hybrid and purebred families

表3 杂交家系的杂种优势率Tab. 3 The heterosis of hybrid families
2.2 各组主要生长性状对体质量的通径分析
通过相关分析, 3个生长性状与体质量的相关系数见表4。4个家系在60 d时的3个生长性状与体质量的相关系数大小依次为壳长>壳高>壳宽; 在127 d时, ZH和HH的3个生长性状与体质量的相关系数大小依次为壳长>壳高>壳宽, HZ的3个生长性状与体质量的相关系数大小依次为壳宽>壳高>壳长, ZZ的3个生长性状与体质量的相关系数大小依次为壳宽>壳长>壳高。
根据通径分析原理, 得到各性状对活体质量的通径系数(表 4), 根据相关系数的组成效应, 可将贝壳各性状与活体质量的相关系数剖分为各性状的直接作用Pi和各性状通过其他性状的间接作用两部分, 即rxiy=Pi+rijPj, 结果见表4。通径系数反映自变量对依变量的直接影响, 结果表明 60 d时, 对于自交家系HH和ZZ壳长对体质量的直接作用最大, 杂交家系ZH和HZ壳长对体质量的直接作用最大。4组壳高壳长和壳宽对体质量的间接作用大于直接作用, 主要是壳高和壳宽对体质量产生的间接作用。在127 d时, 对于自交家系HH壳长对体质量的直接作用最大, ZZ壳宽对体质量的直接作用最大, HH和ZZ壳高、壳长和壳宽对体质量的间接作用大于直接作用, 主要是壳高和壳宽对体质量产生的间接作用。杂交家系ZH壳高对体质量的直接作用最大, ZH的壳高、壳长和壳宽对体质量的直接作用大于间接作用。杂交家系HZ壳宽对体质量的直接作用最大, HZ壳高、壳长和壳宽对体质量的间接作用大于直接作用,主要是壳高和壳长对体质量产生的间接作用。

表4 各性状对活体质量的影响的通径分析Tab. 4 Path analysis of the effects of the economic traits on body weight
2.3 各组主要生长性状对体质量的决定程度
根据单个性状对活体质量的决定系数为Di=Pi2,两个性状对活体质量的共同决定系数Dij=2rijPiPj,计算出贝壳各性状间协同对活体质量的决定系数见表 5。对于杂交家系 ZH, 壳长和壳宽(60 d)的共同决定作用最大, 壳高(127 d)单独决定作用最大;杂交家系 HZ, 壳长(60 d)的单独决定作用最大, 壳宽(127 d)的单独决定作用最大; 自交家系HH壳长和壳宽(60 d)的共同决定作用最大, 壳高和壳长(127 d)共同决定作用最大; 自交家系ZZ壳长(60 d)的单独决定作用最大, 壳宽(127 d)的单独决定作用最大。
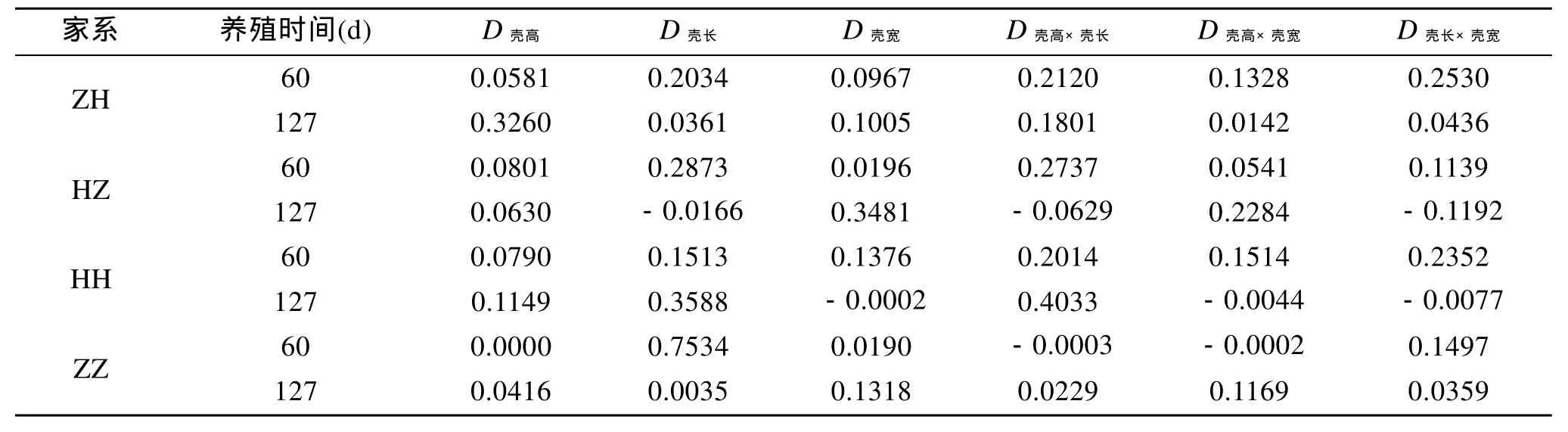
表5 贝壳各性状对活体质量的决定系数Tab. 5 The determinant coefficients of the traits on body weight
2.4 多元回归方程的建立
根据多元相关和通径分析的结果, 以活体质量为依变量, 以对其产生显著影响的其他主要性状为自变量进行了多元回归分析,X1,X2,X3分别是壳长、壳高和壳多元回归方程宽, 建立以壳长、壳高、壳宽为自变量, 活体质量为依变量的多元回归方程, 各组的多元回归方程见表6。结果显示, 各组线性回归关系达到极显著水平(P<0.01)。

表6 多元回归方程Tab. 6 Multiple regression equations
3 讨论
3.1 各组生长比较
海湾扇贝引种养殖的成功极大地推动了中国海水养殖业的发展, 但是, 进入20 世纪90 年代以后,随着海湾扇贝全人工育苗养殖代数的增加, 在生产中逐渐出现一些问题, 例如生长速度减缓、病害频发和商品规格小型化等。产生这些问题的原因与海区环境恶化、养殖结构不合理、病原生物增加等有关,也与连续多代近交可能导致的近交衰退、抗逆能力减弱等引起种质质量下降有关。尽管通过养殖技术和反复引种都能够使上述问题得到一定程度的解决,但是无法从根本上解决问题, 只有借助于遗传育种技术提高其抗逆性和培育新品种, 才能使这些问题从根本上得到解决[19-21]。在养成实验中发现, 紫扇贝和海湾扇贝的杂交子代在壳高、壳长、壳宽和体质量方面和海湾扇贝与紫扇贝相比均表现出较高的生长优势(P<0.01)。在60 d和127 d时, 2个杂交家系的4个经济性状具有一定的杂种优势, 2个杂交家系的壳高、壳长、壳宽和体质量的杂种优势率随着养成时间的增加而逐渐增大。在60 d, 在4个经济性状中, ZH和HZ体质量杂种优势率最大, ZH的壳高和体质量杂种优势率高于HZ, 壳长、壳高和壳宽也表现出一定的杂种优势率, 这表明紫扇贝和海湾扇贝杂交改良扇贝品质是可行的。
3.2 通径分析
方差分析的结果表明, 在各养殖时期, 四个家系的形态性状和体质量均存在显著极差异(P<0.01),根据这一结果来确定影响体质量的优势形态性状是很困难的。在动物育种中, 利用不同性状之间的相关性可为选育工作提供方便。但是简单相关分析不能全面考察变量间的相互关系, 使结果带有一定的片面性。多元回归分析在一定程度上能够消除变量之间的混淆, 但由于偏回归系数带有单位, 使原因对结果的效应不能直接进行比较。生长性状除通过直接作用引起目标性状变化外, 还可通过其他性状以间接作用引起目标性状变化[22]。Wright于1921年提出了通径分析方法, 该方法克服了相关分析与回归分析的不足, 能够真实反映各自变量和依变量的关系, 而且由于通径系数是变量标准化的偏回归系数,从而使原因对结果的效应能直接进行比较[22]。同时通径系数还能区分原因对结果的直接作用和间接作用, 能全面反映原因和结果的相对重要性。由于通径分析把相关系数剖分成两部分, 其中直接影响部分即是通径系数, 且不受其他有关变量的影响, 因此可以通过通径分析来探明两个性状之间真正关系[23]。
通过相关分析, 4个家系在60 d时的3个生长性状与体质量的相关系数大小依次为壳长>壳高>壳宽; 在127 d时, ZH和HH的3个生长性状与体质量的相关系数大小依次为壳长>壳高>壳宽, HZ的 3个生长性状与体质量的相关系数大小依次为壳宽>壳高>壳长, ZZ的3个生长性状与体质量的相关系数大小依次为壳宽>壳高>壳长。在127 d时, ZH壳高和壳长的直接作用大于间接作用, 直接作用最大的是壳高, 在60 d时, 3个生长性状的间接作用均大于直接作用; 其他家系在两个生长时期内 3个生长性状的间接作用均大于直接作用。例如, 在127 d时,ZH的3个生长性状与体质量的相关系数大小依次为壳长>壳高>壳宽, 它的通径系数大小依次为壳高>壳宽>壳长。因此, 采用通径分析不仅可以分析计算出各个自变量与依变量之间的直接关系和间接关系,并且可以发现各个自变量之间存在相互制约关系,才产生了相关系数大而通径系数不一定大的现象。刘小林等[15]在栉孔扇贝的研究中也发现了这一现象。本实验数据所建立的多元回归方程, 经检验, 回归关系极显著, 偏回归关系也极显著, 表明该方程具有较广泛的适用范围和实用价值。
根据本文中通径分析和决定系数分析的结果来看, 以127 d为标准, 在对高活体质量杂交扇贝的选育过程中, ZH家系综合起来看应优先考虑的性状为壳高(直接作用大于间接作用), 其次是壳宽和壳长,壳高、壳宽和壳长均为易测性状, 且壳高对活体质量的通径系数和总的决定系数在所有形态性状中最大,是影响活体质量的最重要的形态性状, 因此, 其可以作为第一目标性状加以选育, 同时, 可把壳宽作为第二、把壳长作为第三目标性状对壳高加以辅助选育, 以达到最佳的选育效果。而HZ家系的3个性状的间接作用大于直接作用, 主要是壳高和壳长对体质量产生的间接作用, 但是比较 3个性状的直接作用发现, 壳宽对体质量的直接作用最大, 可以作为第一目标性状, 同时, 壳高和壳宽做为第二和第三目标性状对壳宽加以辅助选育。
[1]张福绥, 何义朝, 元玲欣, 等. 墨西哥湾扇贝的引种和子一代苗种培育[J]. 海洋与湖沼, 1994, 25(4):272-377.
[2]张福绥. 近现代中国水产养殖业发展回顾与展望[J].世界科技研究与发展, 2003, 25(3): 5-13.
[3]李云福, 刘路伟, 邢光敏, 等. 美国海湾扇贝引种制种及选育技术报告[J]. 河北渔业, 2000, 2: 29-32.
[4]张福绥. 海湾扇贝引种复壮研究[J]. 海洋与湖沼,1997, 28(2): 146-152.
[5]张国范, 刘述锡, 刘晓, 等. 海湾扇贝自交家系的建立和自交效应[J]. 中国水产科学, 2003, 10(6):441-445.
[6]郑怀平, 张国范, 刘晓, 等. 不同贝壳颜色海湾扇贝(Argopecten irradians)家系的建立及生长发育研究[J].海洋与湖沼, 2003, 34(6): 632-638.
[7]郑怀平, 张国范, 刘晓, 等. 海湾扇贝杂交家系与自交家系生长和存活的比较[J]. 水产学报, 2004, 28(3):267-272.
[8]秦艳杰, 刘晓, 张海滨, 等. 海湾扇贝杂交后代的微卫星鉴定[J]. 中国水产科学, 2007, 14(4): 672-677.
[9]Gonzalez M L, Perez M C, Lopez D A. Breeding cycle of the northern scallopArgopecten purpuratus(Lamarck, 1819) in southern Chile [J]. Aquaculture Res,2002, 33: 847-852.
[10]王春德, 刘保忠, 李继强, 刘升平. 紫扇贝与海湾扇贝种间杂交的研究[J]. 海洋科学, 2009, 33(10):83-91.
[11]Wang C, Liu B, Li J, et al. Introduction of the Peruvian scallop and its hybridization with the bay scallop in China[J]. Aquaculture, 2011, 310: 380-387.
[12]佟雪红, 董在杰, 缪为民, 等. 建鲤与黄河鲤的杂交优势研究及主要生长性状的通径分析[J]. 大连水产学院学报, 2007, 22(3): 159-163.
[13]Turker H, Eversole A G. Evaluation of nondestructive method for determining body composition of crayfish[J].Journal of Shellfish Research, 1998, 17(1): 339.
[14]董世瑞, 孔杰, 万初坤, 等. 中国对虾形态性状对体重影响的通径分析[J]. 海洋水产研究, 2007, 28(3):15-22.
[15]刘小林, 常亚青, 相建海, 等. 栉孔扇贝壳尺寸性状对活体重的影响效果分析[J]. 海洋与湖沼, 2002,33(6): 673-678.
[16]刘志刚, 王辉, 孙小真等. 马氏珠母贝经济性状对体重决定效应分析[J]. 广东海洋大学学报, 2007, 27(4):15-20.
[17]何毛贤, 史兼华, 林岳光, 等. 马氏珠母贝生长性状的相关分析[J]. 海洋科学, 2006, 30(11): 1-4.
[18]袁志发, 周敬芋, 郭满才. 决定系数—通径系数的决策指标[J]. 西北农林科技大学学报(自然科学版),2001, 29(5): 131-133.
[19]张福绥, 何义朝, 亓玲欣, 等. 海湾扇贝引种复壮研究[J]. 海洋与湖沼, 1997, 28(2): 146-152.
[20]张国范, 李霞, 薛真福. 我国贝类大规模死亡的原因分析及防治对策[J]. 中国水产, 1999, 9: 34-39.
[21]Chew K K. Global bivalve shellfish introductions [J].World Aquaculture, 1990, 21: 9-22.
[22]李朝霞, 王春德. 海湾扇贝自交与杂交子代的生长比较和通经分析[J]. 中国农学通报2009, 25(8): 282-285.
[23]盛志廉, 吴常信. 数量遗传学[M]. 北京: 中国农业出版社, 1999: 16-26.
Growth and path analyses of hybrid families between the Peruvian and bay scallop
LI Jian-li1, WANG Chun-de1, LI Zhao-xia1, CAI De-quan2, ZHANG Jin-sheng3
(1. Qingdao Agricultural University, Qingdao 266109, China; 2. Weihai Bureau of Ocean and Fisheries, Weihai 264200, China; 3. Xiadian Agricultural Technology Promotion Station, Zhaoyuan 265415, China)
Nov. ,18, 2011
Argopecten purpuratus;Argopecten irradians irradians; hybrid family; heterosis; path analysis
Two inter-specific hybrid families (HZ and ZH) and two inbred families (ZZ and HH) were constructed using the Peruvian scallop (Argopectenpurpuratus) and the bay scallop (Argopecten irradians irradians). Their shell height, shell length, shell width and whole weight were measured on Day 60 and 127 and the major factors affecting their body weight were analyzed. The results showed obvious growth hetereosis in shell height, shell thickness and whole weight (P<0.01). Path analyses showed that shell length was the major factor affecting the whole weight for inbred families. While for the hybrid family HZ, shell length on Day 60 and shell width on Day 127 were the major factors affecting whole weight. In the hybrid family ZH, the interaction between shell length and shell width on Day 60 and the shell height on Day 127 were the major factors affecting whole body weight.Therefore, different parameters should be used in selection of the hybrid families at different stages to ensure the greatest hetereosis.
S917.3
A
1000-3096(2012)08-0015-06
2011-11-18;
2012-04-26
国家自然科学基金项目(31172404); 国家自然科学青年基金(30901101); 农业部引进国际先进农业科学技术(948)项目(2008-Z31,2011-Z27); 青岛市科技计划项目(11-2-1-14-hy)
李建立(1987-), 男, 硕士研究生, 主要从事海洋生物遗传育种学研究, E-mail: lijianli008@126.com; 王春德, 通信作者, 博士, 主要从事海洋生物遗传育种学研究, E-mail: chundewang2007@163.com
梁德海)
