大肠杆菌ATCase抗反馈抑制突变体的构建及其对胞苷积累的影响
2012-01-10方海田周运佼谢希贤
方海田,周运佼,谢希贤,陈 宁
(1. 天津科技大学生物工程学院,天津 300457;2. 宁夏大学农学院,银川 750021)
大肠杆菌ATCase抗反馈抑制突变体的构建及其对胞苷积累的影响
方海田1,2,周运佼1,谢希贤1,陈 宁1
(1. 天津科技大学生物工程学院,天津 300457;2. 宁夏大学农学院,银川 750021)
为解决胞苷生物合成途径中天冬氨酸氨甲酰转移酶受胞苷三磷酸反馈抑制调节的问题,通过对其碱基序列和蛋白质结构分析,利用基因定点突变的方法构建了大肠杆菌的 ATCase突变酶,得到三个突变体:M1(H20L)、M2(K60E)、M3(K94E),并在E. coli DH5α中对融合蛋白进行了表达.酶活测定表明,M1、M2、M3的ATCase酶相对活性都比野生型M0的高,分别为野生型M0的1.10、1.22和1.37倍,且比活力都有不同程度提高.与含野生型pyrBI基因的 M0相比,含突变型基因的 M1、M2和 M3均对 15,mmol/L的 CTP具有强的抗反馈抑制作用,且 M1、M2和M3的抗CTP反馈抑制作用分别是M0的5.4、6.0和8.5倍.最后将各突变质粒转入到E. coli Cyt10(Δcdd)中进行发酵培养,结果表明,与未含突变基因菌株相比,各含突变基因菌株的胞苷积累量均有不同程度的提高,说明 ATCase定点突变使胞苷的合成积累途径得到了不同程度的强化.
大肠杆菌;胞苷;天冬氨酸转氨甲酰酶;定点突变;抗反馈抑制
目前胞苷的生产主要采用发酵法[1],利用基因重组技术构建产胞苷的基因工程菌,通过改变发酵条件积累胞苷,而解除天冬氨酸氨甲酰转移酶的反馈抑制调节作用是发酵法生产胞苷的关键问题之一.天冬氨酸氨甲酰转移酶(aspartate transcarbamylase,ATCase,EC 2.1.3.2)是一种调节酶,由 pyrBI基因编码,催化氨甲酰磷酸和天冬氨酸形成氨甲酰天冬氨酸的反应[2–3].与哺乳动物 ATCase结构不同,大肠杆菌中该酶由两个催化亚基和三个调节亚基组成,催化亚基为相对分子质量3.4×104蛋白质的三聚体,调节亚基为相对分子质量1.7×104蛋白质的二聚体,通过蛋白质桥与调节亚单位互相联结起来[4].pyrB基因编码催化链,pyrI基因编码调节链.胞苷三磷酸(cytidine triphosphate,CTP)是其负调节物,ATP是其正调节物[4–5]. Shepherdson等[6]发现 ATCase的一个最有效的抑制剂是代谢产物CTP,当CTP水平高时,CTP与ATCase结合,降低CTP合成的速度;反之,当细胞内CTP水平低时,CTP从ATCase上解离,加快CTP合成速度.
E. coliK12的ATCase的CTP的可能结合位点分别是 Ile12B、His20B、Lys60B、Val91B、Lys94B[7],如图1所示.据此来设计pyrBI上的突变位点.

图1 CTP在ATCase上的结合位点Fig. 1 Binding sites of CTP in ATCase
为解除 CTP对 ATCase的反馈抑制,本研究针对 ATCase上 CTP的结合位点问题进行研究.在对该酶调控结构域(R-domain)氨基酸序列同源比较的基础上,结合已报道的大肠杆菌全基因组序列和CTP在ATCase上的结合位点,利用合成寡核苷酸引物实现体外定点突变的方法,将高度保守区域进行定点突变[8],期望获得具有较高催化活性且解除产物CTP反馈抑制的酶.最终对比 CTP结合位点突变前后的重组大肠杆菌的 ATCase酶活性和胞苷产量,以考察ATCase定点突变对胞苷积累的影响.
1 材料与方法
1.1 材料
1.1.1 质粒、菌种和培养基
大肠杆菌Escherichia coliK12、E. coli DH5α、E. coli Cyt10(Δcdd)为本实验室保藏;原核表达质粒pSTV28购自TaKaRa公司.
发酵培养基:葡萄糖 16,g,玉米浆 5,mL,豆饼水解液 25,mL,酵母膏 1,g,柠檬酸三钠 2,g,MgSO4·7H2O 5,g,KH2PO42,g,FeSO4·7H2O 100,mg,蒸馏水1,000,mL.
1.1.2 试剂、仪器及分析软件
氨甲酰基磷酸盐、L–天冬氨酸、胞苷三磷酸,Sigma公司;Pfu聚合酶,Fermentas公司;基因定点突变试剂盒、限制性内切酶BamHI、HindⅢ、DpnI、T4,DNA 连接酶、DNA Marker DL10000,大连TaKaRa公司;硅胶模型TM基因组 DNA 提纯试剂盒、TianzTMPCR产物(DNA片段)纯化试剂盒、硅胶模型TM质粒 DNA小量提纯试剂盒,北京博迈德公司;Easy Protein Quantitative Kit,北京全式金公司.
PCR仪,Bio-Rad公司;SBA–40B生物传感仪,山东生物科学研究所.DNA 序列分析比较采用DNAMAN 2.0软件,引物设计采用Primer Premier 5软件,电泳图分析采用 Imagemaster TotalLab v1.00软件.
1.2 实验方法
1.2.1 重组大肠杆菌表达质粒的构建
根据 NCBI上检索到的大肠杆菌 K12标准株MG1655的 ATCase基因 pyrBI的核苷酸序列(GenBank:U00096)设计引物,用于 ATCase基因的克隆和定点突变.以大肠杆菌K12基因组DNA为模板,用引物P1和P2将pyrBI基因扩增出来,以常规分子克隆手段连入pSTV28载体中,所含抗性标记为Cmr,得到重组质粒.


1.2.2 重组大肠杆菌表达质粒的定点突变
利用基因定点突变试剂盒(Site-directed Gene Mutagenesis Kit)用于点突变[9].对所构建的重组质粒进行 PCR.从基因组中克隆出目的基因片段并分别引入突变位点进行质粒 PCR.将定点突变后的 pyrBI送华大基因公司进行核苷酸测序.
基因定点突变反应体系:Nuclease-Free Water,41,µL;Reaction Buffer(10×),5,µL;引物(10,µmol/L each),2,µL;dNTP Mix(10,mmol/L each),1,µL;待突变模板质粒(0.5,µg),0.5,µL;Pfu DNA Polymerase,0.5,µL.PCR 反应条件:预变性 95,℃ 3,min;变性95,℃ 30,s;退火 60,℃ 30,s;延伸 72,℃ 4.5,min;延伸、补全 72,℃ 10,min;4,℃保持;共 35个循环.PCR反应后,加入 1,µL DpnI,混匀后 37,℃孵育 1,h.DpnI消化完毕后可以直接用于转化.
转化、挑克隆鉴定:在100,µL DH5α感受态细菌中加入5~10,µL经过DpnI消化后的突变产物,冰浴20,min,42,℃热击 60,s,冰浴 2,min 后加入 890,µL SOC 培养基,37,℃、180,r/min复苏 1,h.在涂板前5,000,r/min离心 1,min,全部均匀地涂布到含有氯霉素的 LB平板上,37,℃培养过夜.进行菌落 PCR,然后先挑单个克隆提取质粒,酶切验证.取 3~5个酶切鉴定正确的克隆测序,以最终确认得到的克隆是否是预期的突变克隆.
1.2.3 野生型及突变型酶基因的诱导表达及酶活性测定
将重组菌于 37,℃培养至 A600为 0.6~1.0,加入0.1,mmol/L IPTG,收集菌体,经超声破碎后取上清液,以 12% SDS-PAGE鉴定,并测定 ATCase酶活.酶活测定反应混合物:50,mmol/L Tris-HCl(pH 7.0),15,mmol/L L–天冬氨酸,8,mmol/L Na2-CP,加ATCase酶液补足1,mL.28,℃保温30,min,加热煮沸终止反应,摇匀后 466,nm 比色.酶活力单位定义为:在 28,℃条件下,每分钟催化生成 1,µmol 产物(氨甲酰天冬氨酸)为一个酶活力单位(U).酶的比活力为酶活力除以蛋白质质量分数所得值,蛋白质质量分数的检测参照 Bradford法[10],利用 Easy Protein Quantitative Kit直接测定.
1.2.4 CTP对 ATCase活性的影响及含突变基因重组子对CTP的抗反馈抑制作用的测定
在测定反应体系中分别加入终浓度为 0、0.5、1、2、5、10、15,mmol/L 的 CTP,按标准酶活测定方法测定酶活.以未添加 CTP时酶活力设为 100%,计算相对活性.
1.2.5 突变体的发酵培养及对胞苷积累量的测定
将得到的突变质粒分别导入到 Escherichia coli Cyt10(Δcdd)中,在 37,℃、200,r/min 条件下进行发酵培养.发酵培养 40,h后结束发酵,采用 HPLC测定反应生成的胞苷.
2 结果与分析
2.1 野生型与突变体大肠杆菌融合蛋白表达质粒的构建及鉴定
分别将野生型pyrBI基因和突变型pyrBI基因连接到表达载体 pSTV28上,导入到 DH5α中.然后将大肠杆菌重组质粒转化子进行验证,对重组菌进行菌落 PCR,将菌落 PCR 产物用琼脂糖凝胶电泳检测.根据菌落 PCR的筛选结果,从每种重组质粒的单菌落中挑取菌体培养,取适量培养液,用博迈德公司的质粒 DNA小量提取试剂盒提取质粒,将提纯的质粒分别用 BamHI和 HindⅢ进行单酶切反应,同时进行双酶切反应.对酶切反应液用琼脂糖凝胶电泳检测反应产物,结果如图2所示.

图2 突变体重组质粒的双酶切验证Fig. 2 DNA electrophoresis of identification of recombined plasmid by digestion with BamHI and HindⅢ
将突变体质粒送北京华大基因公司测序.对所得基因进行测序和序列分析表明,ATCase上的His20B已突变为 Leu20B,Lys60B突变为 Glu60B,Lys94B突变为Glu94B突变的基因,所得ATCase基因序列与设计的突变相符.
2.2 野生型与突变体大肠杆菌 ATCase的诱导表达及酶活分析
将重组大肠杆菌M0、M1、M2、M3分别培养,用0.1,mmol/L IPTG诱导表达,然后取菌样提取粗酶液.进一步测定 ATCase酶活进行验证可知,重组的基因工程菌经过诱导表达后可表达出相对活性较高的 ATCase,明显优于未转化重组质粒的大肠杆菌,结果如图3所示.
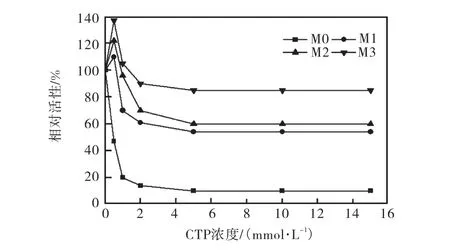
图3 不同CTP浓度下野生型与突变型ATCase的反馈抑制作用Fig. 3 Feedback inhibition of ATCase by CTP of the original and mutant on different CTP concentration
突变体的酶活均有升高,分别是野生型的 1.10、1.22、1.37倍.不同突变体的 ATCase的比活力及胞苷产量见表1.突变体M1、M2和M3的比活力分别比野生型M0提高11.2%、20.6%和35.1%.
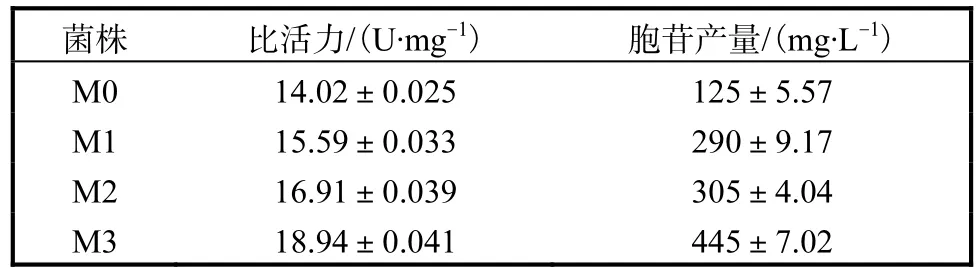
表1 不同突变体的ATCase的比活力及胞苷产量Tab. 1 ATCase specific activities and cytidine production of different mutants
2.3 野生型和突变型酶蛋白抗CTP反馈抑制比较
对于野生型酶,当酶活反应体系中的 CTP的浓度达到 0.5,mmol/L时,就会产生反馈抑制而导致酶活力下降.为了构建胞苷的高产菌株,必须使酶对CTP的抗反馈抑制作用达到一定程度.如图3所示,野生型 ATCase在 CTP浓度达到 0.5,mmol/L时,ATCase活性仅存47%;在CTP浓度达到15,mmol/L时,M0的 ATCase活性仅存 10%,突变体 M3的ATCase活性仍有 85%以上,而突变体M1与 M2的ATCase活性分别为54%和60%;M1、M2和M3与野生型M0相比,M1、M2和M3的抗CTP反馈抑制作用分别是M0的5.4、6.0和8.5倍,对CTP的抗反馈抑制作用也有一定程度的提高.
2.4 野生型与含突变基因重组菌对胞苷产量的影响
对比摇瓶发酵结果,以考察含突变后的 pyrBI基因的各突变体大肠杆菌是否去除了 CTP的反馈抑制问题.对比 CTP结合位点突变前后的重组大肠杆菌M0、M1、M2和M3的胞苷产量.
各突变体菌株产生的胞苷分别为(290±9.17)、(305±4.04)、(445±7.02)mg/L,分别是突变前菌株M0(125±5.57)mg/L 的 2.32、2.44、3.56倍.实验结果表明,针对 CTP在 ATCase上的结合位点进行定点突变有显著效果,说明突变 pyrBI基因所表达的ATCase的反馈抑制问题得到了改善,强化了胞苷合成途径.
3 讨 论
ATCase酶催化嘧啶从头合成途径中的第二步反应,是重要的限速酶.为了获得抗代谢产物 CTP反馈抑制且保持较高酶催化活性的酶蛋白,本研究采用基因定点突变(site-directed gene mutagenesis)的方法,有目的地构建突变体,通过对野生型酶蛋白和突变型酶蛋白性质的研究,筛选能较好地保持 ATCase酶的活性且具有抗反馈抑制性质的突变体.
由于CTP对ATCase的反馈抑制与调节亚基相关,所以要寻找起关键作用的 CTP结合位点进行突变,得到 3个突变体,从而实现对酶蛋白的精细改造,期望可以解除反馈抑制作用.根据几种相关微生物 ATCase调节亚基氨基酸序列多重比较的结果,结合已有的研究结果[11],利用扩增出的未突变和突变pyrBI基因及 pSTV28质粒,成功地构建了用于大肠杆菌表达的重组质粒,并将其转化入 E. coli Cyt10(Δcdd).成功构建了 M0(野生型)、M1(H20L)、M2(K60E)、M3(K94E).酶学性质研究表明:几种突变酶比野生型酶具有较高的抗 CTP反馈抑制的能力,且 ATCase活性均有一定程度的提高.pSTV28质粒属于低拷贝质粒,为避免因拷贝数高导致的高表达量对发酵目标产物产率的对应关系产生影响,故选择低拷贝数质粒以消除这种影响.表明这些位点可能与酶蛋白与 CTP的结合或酶的多聚化有关;突变体酶活性的提高可能与酶易与底物结合有关;也可能与因突变导致氨基酸发生改变从而减弱 CTP在酶上的结合所需的作用力如氢键、离子键及疏水作用力有关.
本研究在酶学性质的体外研究基础上,通过含突变酶基因的重组子进行酶的体内活性研究.野生菌中,当 CTP浓度达到 5,mmol/L时,会导致酶活力降低.而突变后的酶蛋白不易与 CTP发生结合,酶活性不受影响,当CTP浓度为15,mmol/L时,含突变基因的菌株都可以旺盛地生长.突变后,各突变体的酶比活力均有不同程度的提高.总之,本研究成功地构建了既部分解除了 CTP的反馈抑制又保持了酶催化活性的M1、M2和M3三个酶突变体.
野生型 M0在发酵培养中胞苷产量和 ATCase抗 CTP反馈抑制效应显著低于突变型 M1、M2和M3,表明突变pyrBI基因所表达的ATCase的受CTP反馈抑制的问题得到了改善,在未经优化的条件下通过发酵培养,使得胞苷的积累量有了一定程度的提高.但是,胞苷的产量并没有大量积累,可能是胞苷合成途径中还存在其他负调控因素,如 UMP对CPSase和ATCase的反馈抑制,CTP对CTP合成酶的反馈抑制及阻遏等[12–13].解除 CTP反馈抑制突变菌株的成功构建为发酵法生产胞苷过程的深入研究提供了很好的基础.目前,突变体正进一步用于构建胞苷的高产菌株的研究中.
[1]苏静,黄静,谢希贤,等. 枯草芽孢杆菌 cdd基因敲除及对胞苷发酵的影响[J]. 生物技术通讯,2010,21(1):39-42.
[2]方海田,谢希贤,徐庆阳,等. 微生物发酵法生产胞嘧啶核苷的研究进展[J]. 发酵科技通讯,2010,39(3):48-51.
[3]Nelson D L,Cox M M. Lehninger Priciples of Biochemistry[M]. 3rd ed. New York:Worth Publishers,2000:818-868.
[4]Simmer J P,Kelly R E,Scully J L,et al. Mammalian aspartate transcarbamylase(ATCase):Sequence of the ATCase domain and interdomain linker in the CAD multifunctional polypeptide and properties of the isolated domain [J]. Proceedings of National Academy of Sciences of United States of America,1989,86(12):4382-4386.
[5]Rabinowitz J D,Hsiao J J,Gryncel K R,et al. Dissecting enzyme regulation by multiple allosteric effectors:nucleotide regulation of aspartate transcarbamoylase[J].Biochemistry,2008,47(21):5881–5888.
[6]Shepherdson M,Pardee A B. Production and crystallization of aspartate transcarbamylase[J]. The Journal of Biological Chemistry,1960,235(11):3233-3237.
[7]Heng S,Stieglitz K A,Eldo J,et al. T-state inhibitors of E. coli aspartate transcarbamoylase that prevent the allosteric transition[J]. Biochemistry,2006,45(33):10062-10071.
[8]张蓓. 代谢工程[M]. 天津:天津大学出版社,2003.
[9]徐芳,姚泉洪,熊爱生,等. 重叠延伸 PCR技术及其在基因工程上的应用[J]. 分子植物育种,2006,4(5):747-750.
[10]Bradford M M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding[J]. Analytical Biochemistry,1976,72:248-254.
[11]张远富,徐莹,Van de Casteele M,等. 嗜冷弧菌ATCase的酶学特性[J]. 微生物学通报,1999,26(1):41-44.
[12]Berg J,Tymoczko J L,Stryer L. Biochemistry[M]. 6th ed. New York:W. H. Freeman Company,2002.
[13]苏静,邓培生,谢希贤,等. 基于 cdd基因敲除和嘧啶操纵子转移的胞苷产生菌的研究[J]. 天津科技大学学报,2010,25(5):1-5.
Construction of ATCase Mutants with Feedback Inhibition Resistance and their Effect on the Cytidine Production in E. coli
FANG Haitian1,2,ZHOU Yunjiao1,XIE Xixian1,CHEN Ning1
(1. College of Biotechnology,Tianjin University of Science & Technology,Tianjin 300457,China;2. College of Agriculture,Ningxia University,Yinchuan 750021,China)
In order to obtainE. coliATCase mutants with high catalytic activities and feedback inhibition resistance,sitedirected mutagenesis,homologial analysis of the related amino acid sequences of ATCaseand PCR were used to construct three mutants:M1(H20L),M2(K60E),M3(K94E). First,pyrBI gene of M0(wild type)was ligated to plasmid pSTV28 vector,and site-directed mutagenesis was carried out by PCR. The fusion proteins were expressed inE. coliDH5α,and the erzyme activity analysis showed that the original ATCase relative activity of M0 not only remained but also increased by 1.10-fold,1.22-fold and 1.37-fold respectively. Their specific activities were also increased. Measuring of the feedback inhibition resistance of the enzyme showed thatE. coliDH5αstrains containing the mutation gene had a strong feedback inhibition resistance against 15 mmol/L of CTP concentration. The enzymic activities found in M1,M2 and M3 strains were 5.4-,6.0- and 8.5-fold of M0 in CTP feedback inhibition resistance of ATCase regulation. Finally,all mutant plasmids were transformed intoE. coliCyt10(Δcdd). After fermentation,ATCase synthesis of site-directed mutagenesis indicated that the accumulation of cytidine pathway got strengthened.
E. coli;cytidine;aspartate transcarbamylase;site-directed mutagenesis;feedback inhibition resistance
Q784
A
1672-6510(2012)04-0012-05
2011–11–30;
2012–02–05
天津科技大学科学研究基金资助项目(20100211)
方海田(1978—),男,内蒙古人,博士研究生;通信作者:陈 宁,教授,ningch@tust.edu.cn.
郎婧
