基于水温垂直结构的西北太平洋柔鱼栖息地模型构建*
2012-01-10陈新军
陈新军,陈 峰,高 峰,雷 林
(1.上海海洋大学海洋科学学院;2.上海海洋大学大洋生物资源开发和利用上海市高校重点实验室;3.上海海洋大学大洋渔业资源可持续开发省部共建教育部重点实验室,上海201306)
柔鱼(Ommastrephes bartramii)作为1种大洋性种类,广泛分布在太平洋的亚热带和温带海域,资源极为丰富[1]。该资源于1970年代初期首先由日本渔船开发,此后韩国、中国(包括台湾省)等国家和地区对其进行大规模商业性开发利用。其中西北太平洋柔鱼冬春生产卵群体是中国大陆鱿钓渔船传统的捕捞对象,作业时间从每年的6月持续至11月,8~10月为渔汛旺期,该渔场的渔获量约占其年产量的70%~80%[2]。国内外学者对北太平洋柔鱼种群结构[3]、年龄生产[4-6]、繁殖[7-9]、洄游方式[10-11]、柔鱼及渔场分布与海洋环境关系[12-20]等方面做了大量的研究。由于柔鱼对海洋环境变化较敏感,并且具有昼夜垂直洄游的习性[10],因此,柔鱼中心渔场的形成不仅仅受到海洋表温(Sea Surface Temperature,SST)、锋面、海流等海洋环境要素的影响,同时还受到水温垂直结构的影响[21]。
栖息地指数模型(Habitat Suitability Index,HSI)最早由美国地理调查局国家湿地研究中心鱼类与野生生物署于1980年提出,被用来描述野生动物对周围栖息地环境适应程度[22]。近年来栖息地理论和方法被逐渐应用于渔业资源、渔场的研究中[23-25]。Song和Zhou[23]利用不同水层温度梯度建立栖息地模型分析印度洋大眼金枪鱼分布;冯波等[26]利用分位数回归方法建立基于各水层水温差和溶解氧差的印度洋大眼金枪鱼栖息地模型;郭爱等[27]利用水温垂直结构研究了中西太平洋鲣鱼的栖息地环境;Chen等[28]根据海洋表层环境因子建立了西北太平洋柔鱼栖息地模型,没有涉及到深层水温;Tian等在其研究中已考虑到了深层水温对西北太平洋柔鱼渔场的影响[24],但并没有涉及到各水层水温梯度对柔鱼渔场的影响。因此,本文主要根据1998-2005年西北太平洋柔鱼生产资料,以及各水层温度、水温垂直梯度建立基于水温垂直结构的栖息地模型,分析探讨不同方法所建立的栖息地模型与渔场分布关系,并进行实例验证,为实际渔业生产和管理提供决策依据。
1 材料与方法
1.1 材料来源
(1)生产数据 采用1998—2005年8~10月中国大陆西北太平洋柔鱼生产数据,主要作业海域为150°E~165°E,38°N~46°N,数据字段包括日期、船名、经度、纬度、产量、作业次数。其时间分辨率为天,将0.5(°)×0.5(°)表示为1个渔区,统计1个渔区内作业次数(单位:d)和产量(单位:t)。
(2)海洋环境数据 表层水温(SST)和深层水温数据(50、200和300 m水温),均来自哥伦比亚环境数据(CARTON-GIESE Simple Ocean Data Assimilation,http://iridl.ldeo.columbia.edu/SOURCES/.CARTON-GIESE/SODA/),时间跨度为1998—2005年,月份为8~10月,空间分辨率为0.5(°)×0.5(°)。
1.2 分析方法
(1)以SST、各水层温度和0~50 m、200~300 m温度梯度[29]为基础,统计1998—2004年8~10月0.5(°)×0.5(°)渔区内产量和作业次数。
(2)作业渔船的分布(即作业次数)反映了鱼类偏好或者捕捞几率的分布[30],因此,作业次数可用来表征柔鱼中心渔场的分布。假定最高作业次数为渔业资源丰度较高的海域,指定其栖息地指数HSI为1;作业次数为0时,则认为是鱼类资源丰度较低的海域,指定其HSI为0[31]。栖息地指数大于0.6的海域,认为是鱼类资源较为适宜的栖息海域[28]。通过上述对柔鱼渔场与各水层水温的分析,获得渔场适宜的水温指标。以作业次数为基础建立各水层水温的适应性指数(Suitability Index,SI),利用下列公式计算单因素适应性指数:

式中:SIi为单因素适应性指数;Neti为i渔区作业次数;Netmax为各渔区中的最高作业次数。
(3)利用一元非线性回归法建立SST、50 m水温(T50)、200 m水温(T200)、300 m水温(T300)、0~50 m温度梯度(G0-50)和200~300 m温度梯度(G200-300)与SI之间的关系模型。通过此模型将SST、T50、T200、T300、G0-50和G200-300与SI离散变量关系转化为连续随机变量关系。
(4)利用目前较为普遍的算术平均法(Arithmetic Mean Model,AMM)[32]和几何平均法(Geometric Mean Model,GMM)[33]建立栖息地指数模型。
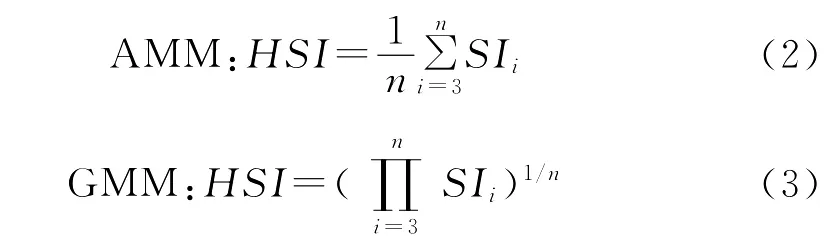
式中,SIi为第i个环境变量的适应性指数;n为模型中的参数个数。
利用式(2)和式(3)不同海洋环境因子的组合,获得综合栖息地指数。根据综合栖息地指数与1998—2004年8~10月作业次数和产量的关系,选择其中HSI大于0.6以上,作业次数和产量比重最高的海洋环境因子组合作为最优模型的环境因子。
(5)以CPUE(单位:t/d)指标作为柔鱼资源丰度,计算公式如下:

式中:CPUE为渔区i的柔鱼资源丰度(t/d);Ci为渔区i的产量(单位:t);Fi为渔区i的作用次数(单位:d)。
1.3 HSI模型的验证
利用2005年8~10月生产数据进行实例验证,以验证基于水温垂直结构的栖息地指数模型(HSI),探讨以此模型来预测柔鱼中心渔场和潜在渔场的可行性。
2 结果
2.1 渔场与各水层温度分布
8月,作业次数主要分布在SST、T50、T200、T300、G0-50和G200-300分别为17~21℃、7~10℃、3~6℃、3~5℃、0.15~0.35℃/m和-0.01~0.01℃/m的海区,分别占总作业次数的75.6、43.5、77.2、85.8、86.1和90.0%(见图1);9月,作业次数主要分布在SST、T50、T200、T300、G0-50和G200-300分别为16~19℃、7~10℃、3~5℃、3~5℃、0.12~0.32℃/m和-0.01~0.015℃/m海区,分别占总作业次数的79.9、41.5、70.9、98.7、70.7和94.7%(见图2);10月,作业次数主要分布在SST、T50、T200、T300、G0-50和G200-300分别为14~16℃、10~13℃、4~6℃、4~5℃、0.04~0.16℃/m和-0.008~0.008℃/m海区,分别占总作业次数的66.3%、64.1%、78.7%、85.8%、82.5%和92.3%(见图3)。同时,分别建立了基于SST、T50、T200、T300、G0-50和G200-300的适应性指数模型(见表1),各模型统计显著(P<0.01)。
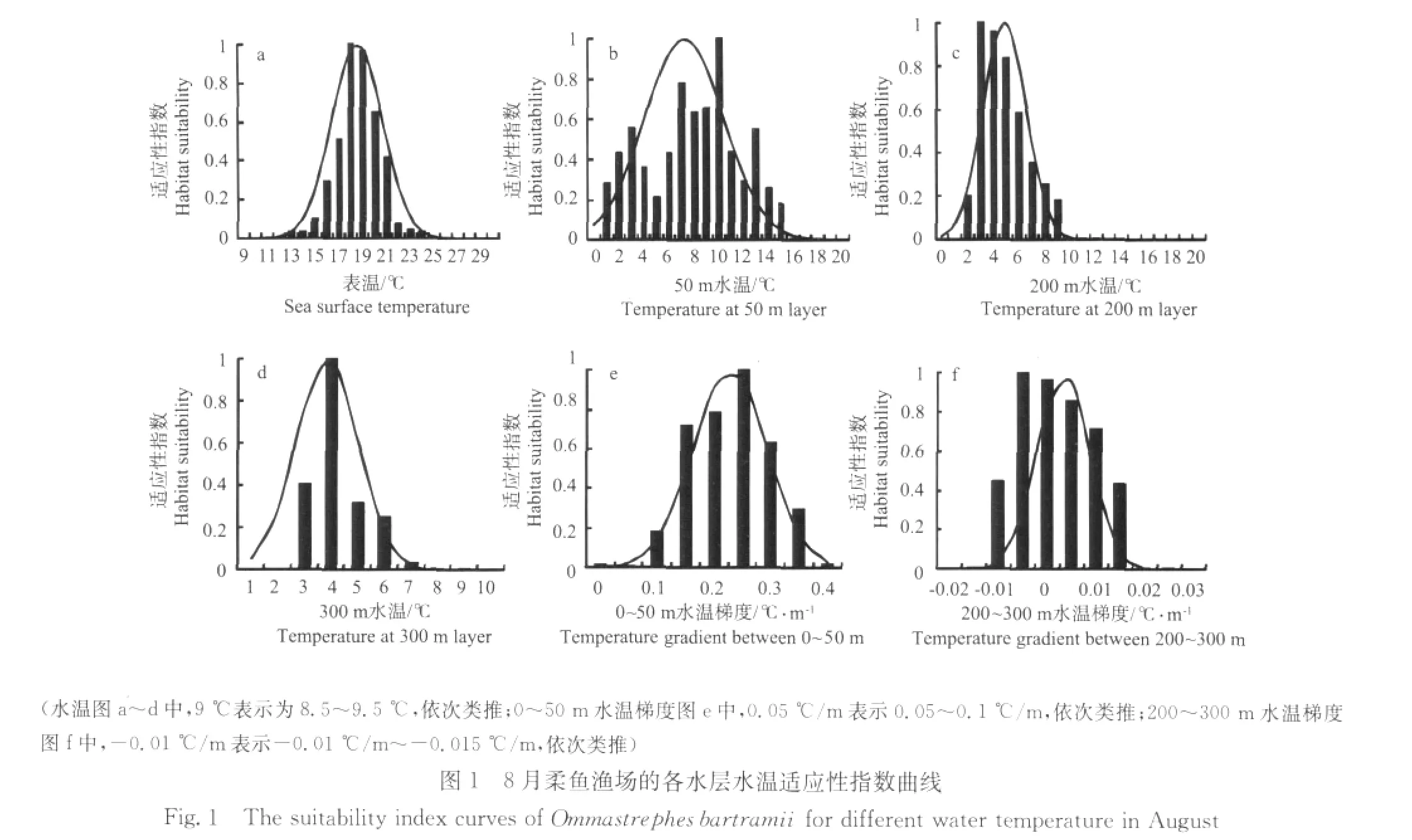


图3 10月柔鱼渔场的各水层水温适应性指数曲线Fig.3 The suitability index curves of Ommastrephes bartramii for different water temperature in October
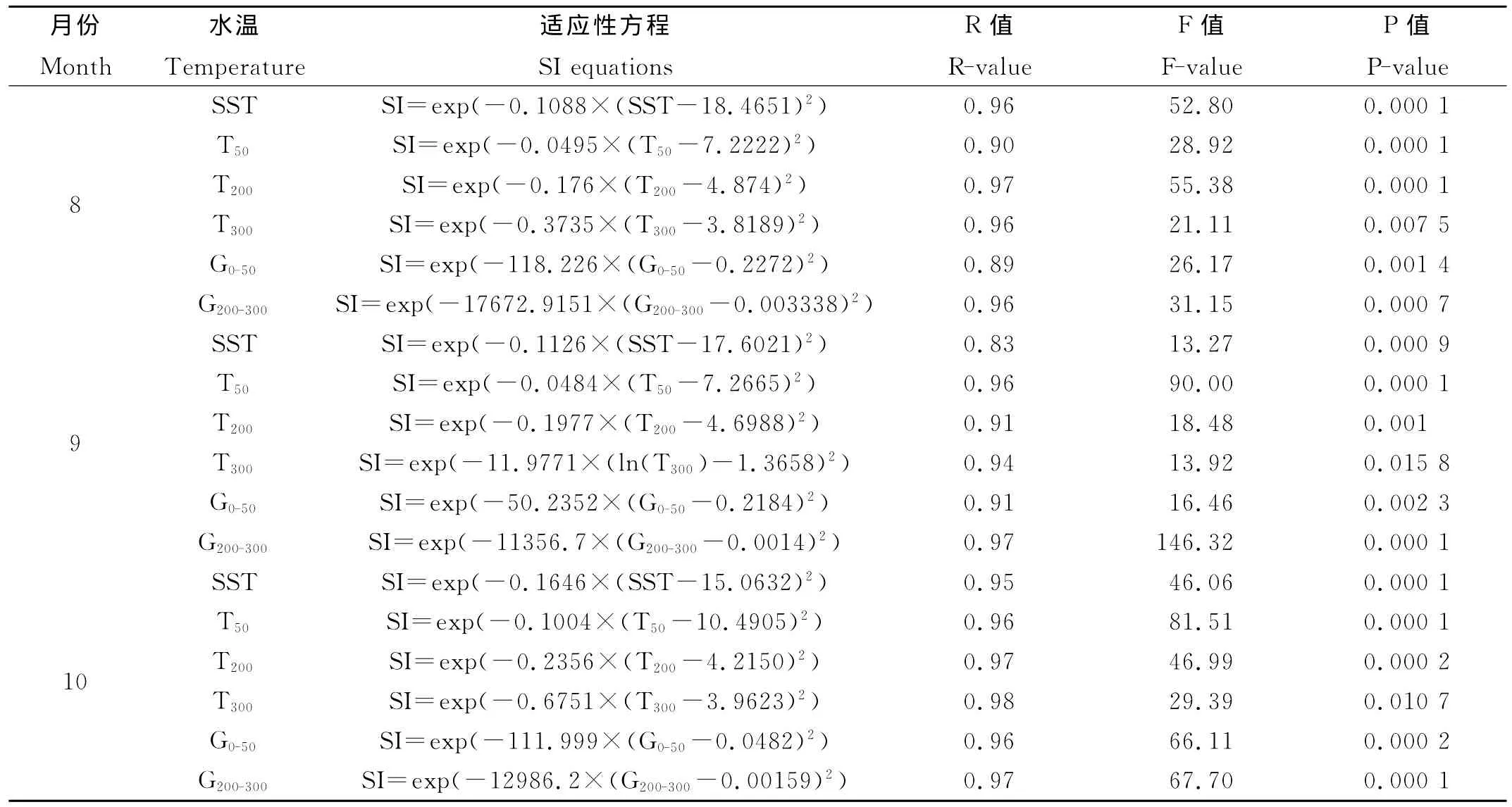
表1 单因素因子的适应性指数方程Table 1 The suitability index equations based on single factors
2.2 HSI模型分析
根据不同环境因子组合获得的综合栖息地指数与HSI>0.6时作业次数和产量比重如表2。相同环境因子条件下,AMM模型中,产量和作业次数比重达到70%以上;而GMM模型中,产量和作业次数比重达到60%以上,可见AMM建立的栖息地指数模型明显优于GMM。利用SST、G0-50、T200和T3004个因子和利用SST、T300和T3003个因子建立的栖息地指数大于0.6时,作业次数和产量在各模型中均比例较高;但综合考虑到0~50 m内水温变动较为剧烈,因此,选取SST、0~50 m水温梯度、200 m水温和300 m水温建立栖息地指数模型。

表2 1998—2004年8~10月栖息地指数在0.6以上情况下作业产量和作业次数所占的比重Table 2 The percentages of catch and fishing effort of Ommastrephes bartramii during August to October from 1998 to 2004 on the basis of values above 0.6 estimated from the AMM and GMM model
根据SST、0~50 m水温梯度、200 m和300 m水温分别计算8~10月适应性指数,然后利用AMM和GMM方法获得柔鱼综合栖息地指数HSI(见图4)。HSI>0.6时,AMM的产量和作业次数比重分别占83.4%和80.9%,CPUE均2.1 t/d以上;GMM的产量和作业次数比重分别占73.5%和69.6%,CPUE均2.3 t/d以上。HSI<0.4,AMM的产量和作业次数比重分别占4.2%和3.9%,CPUE为1.5~2.3 t/d;GMM的产量和作业次数比重分别占10.1%和9.6%,CPUE为1.4~2.4 t/d。分析表明,AMM和GMM均能预测柔鱼中心渔场,预测精度达到70%,但AMM的产量和作业次数比重稍高于GMM,AMM模型稍优于GMM。

图4 1998—2004年8~10月柔鱼产量(a)、作业次数(b),CPUE(c)与HSI关系Fig.4 The relationship between catch(a),fishing effort(b),CPUE(c)of Ommastrephes bartramii and HSI during August to October from 1998 to 2004
2.3 HSI模型验证
根据AMM建立HSI模型,利用2005年8~10月进行实例验证,分别计算各月的HSI值,并与实际作业次数进行比较(见图5和表3)。分析表明,8~10月HSI>0.6海域总的捕捞产量和作业次数比重分别为85.6%和82.5%,CPUE为3.2~4.2 t/d。
8月,HSI>0.6分布在150°E~155°E、40°N~43°N,155°E~157°E、42°N~43°N,157°E~160°E、38°N~44°N和161°E~164°E、38°N~41°N带状海区,作业渔场主要分布在150°E~157°E、40°N~44°N海区,中心渔场HSI>0.6(见图5a);9月,HSI>0.6主要分布在151°E~157°E、41°N~45°N和158°E~165°E、40°N~44°N两海区,而柔鱼渔场主要分布在152°E~157°E、42°N~44°N海区,中心渔场HSI>0.6(图5b);10月HSI>0.6主要分布在153°E~158°E、41°N~46°N和160°E~165°E、40°N~45°N海区,柔鱼渔场主要分布在152°E~157°E、41°N~45°N海区,中心渔场HSI>0.6(见图5c)。
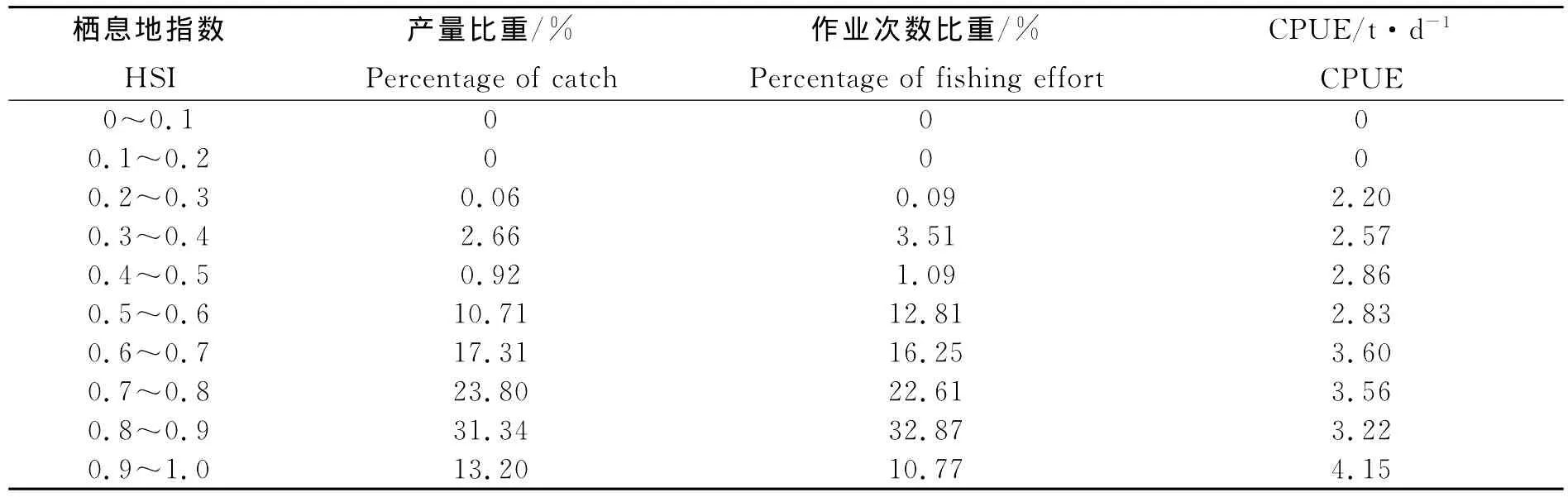
表3 基于HSI的2005年8~10月产量、作业次数和CPUE统计分布Table 3 The catch,fishing effort and CPUE of Ommastrephes bartramii from August to October in 2005 on the basis of values estimated from the HSI model

图5 2005年8~10月柔鱼渔场与基于AMM的HSI分布Fig.5 The relation of fishing ground distribution of Ommastrephes bartramii to HSI based on AMM from August to October in 2005
3 结论与分析
(1)渔场与水温垂直结构关系。柔鱼作为一种短生命周期的种类,其资源变动、渔场分布对海洋环境变化较敏感[34]。SST作为一种找柔鱼中心渔场的重要指标,对柔鱼渔场的变动起着最重要的作用。8~10月柔鱼渔场主要分布在黑潮前锋附近及表温等温线密集的冷暖水交汇区,深层冷暖水交汇处暖水一侧或暖水舌突出处[35]。适宜的水温结构有利于柔鱼的集群以及渔场的形成(图1~3)。如2009年,深层(100m)冷水入侵传统渔场(42°N~46°N,150°E~165°E),使得柔鱼旺汛期作业渔场水温比往年偏低,鱿钓产量大幅下降且渔获率较低[36]。由此可见,表温和深层水温均影响柔鱼资源及渔场分布。因此,有必要在建模的过程中引入深层水温作为单因子,建立水温垂直结构的栖息地指数模型,研究其与渔场分布的关系。同时研究中也发现,50 m以内受到混合层影响,水温变动较大,分布偏度较大。因此,在建模过程引入了0~50 m水温梯度变量,模型预测较好,进一步体现出0~50 m水温梯度的重要性。
(2)HSI模型的讨论。把作业次数作为捕捞努力量,是拟合适应性指数(SI)的较好指标。Tian等[24]曾利用基于CPUE的HSI预测柔鱼的最适栖息海域,由于受到作业渔船、作业技术以及其他海洋环境因素的影响,其预测效果稍低于基于捕捞努力量的HSI模型。适应性指数(SI)表明,柔鱼渔业捕捞努力量即作业次数与各层水温、水温梯度存在着正态分布关系(P<0.05),这一关系也在其它鱼类资源中得到证实[37-38]。
AMM和GMM表明,HSI大于0.6时,8~10月产量和作业次数比重均达到80%,随着HSI增大,产量与作业次数比重不断增大(见图4a,b)。CPUE均在2.1 t/d以上,波动较小,但HSI低于0.4时,AMM的CPUE为1.5~2.3 t/d,而GMM的CPUE为1.4~2.4 t/d,波动明显较大(见图4c)。综合分析认为,在预测西北太平洋柔鱼中心渔场时,AMM稍优于GMM,并且AMM在2005年8~10月作业渔场预测分析中得到了一定的验证。陈新军等[25]利用表温及表温梯度建立的栖息地模型预测渔汛期渔场预测精度也达到80%以上(HSI>0.6)。然而柔鱼渔场不仅仅受到水温的影响,同时也受到海流、叶绿素浓度、海面高度等影响,因此,在以后的分析中需要进一步考虑多环境因子(如表层水温,叶绿素和Argo深层环境因子等)对柔鱼渔场进行动态分析,以实现更加准确地预报中心渔场。
[1] Roper C,Sweeney M,Nauen C.FAO Species Catalogue.Vol.3,Cephalopods of the World.An annotated and illustrated catalogue of species of interest to fisheries[J].FAO Fish Synop,1984,125(3):1-277.
[2] 陈新军,钱卫国,刘必林,等.利用衰减模型评估柔鱼西部冬春生群体资源[J].海洋湖沼通报,2008,2:130-140.
[3] Yatsu A,Tanaka H,Mori J.Population Structure of the Neon Flying Squid,Ommastrephes bartramii,in the North Pacific O-cean[C]∥Contributed Papers to International Symposium on Large Pelagic Squids,Tokyo:Japan Marine Fishery Resources Research Center.1998:31-48.
[4] Yatsu A,Midorikawa S,Shimada T,et al.Age and growth of the neon flying squid,Ommastrephes bartramii,in the North Pacific Ocean[J].Fisheries Research,1997,29(3):257-270.
[5] Yatsu A,Mori J.Early growth of the autumn cohort of neon flying squid,Ommastrephes bartramii,in the North Pacific Ocean[J].Fisheries research,2000,45(2):189-194.
[6] 黄洪亮,郑元甲,程家骅.北太平洋海区柔鱼生物学特征研究[J].海洋渔业,2003,25(3):126-129.
[7] Hayase S.Distribution of spawning grounds of flying squid,Ommastrephes bartramii,in the North Pacific Ocean[J].Japan Agricultural Research Quarterly,1995,29:65-65.
[8] Watanabe H,Kubodera T,Ichii T,et al.Feeding habits of neon flying squid Ommastrephes bartramii in the transitional region of the central North Pacific[J].Marine Ecology Progress Series,2004,266:173-184.
[9] Yatsu A,Mochioka N,Morishita K,et al.Strontium/calcium ratios in statoliths of the neon flying squid,Ommastrephes bartramii(Cephalopoda),in the North Pacific Ocean[J].Marine Biology,1998,131(2):275-282.
[10] Murata M,Nakamura Y.Seasonal migration and diel vertical migration of the neon flying squid,Ommastrephes bartramii in the North Pacific[R].[s.l.]:Contributed papers to International Symposium on Large Pelagic Squids,July 18-19,1996,for JAMARC's 25th anniversary of its foundation Japan Marine Fishery Resources Research Center,1998:13-30.
[11] Ichii T,Mahapatra K,Sakai M,Okada Y.Life history of the neon flying squid:effect of the oceanographic regime in the North Pacific Ocean[J].Marine Ecology Progress Series,2009,378:1-11.
[12] Gong Y,Kim Y S,An D H.Abundance of neon flying squid in relation to oceanographic conditions in the North Pacific[J].International North Pacific Fisheries Commission Bulletin,1993,53:191-204.
[13] Ichii T,Mahapatra K,Sakai M,et al.Differing body size between the autumn and the winter-spring cohorts of neon flying squid(Ommastrephes bartramii)related to the oceanographic regime in the North Pacific:a hypothesis[J].Fisheries oceanography,2004,13(5):295-309.
[14] Murata M,Ishii M,Shingu C.Seasonal changes in location and water temperature of the fishing grounds by jigging fishery for flying squid,Ommastrephes bartramii(LeSueur),with some considerations on migration and occurrence of the fishing ground[J].Bull Hokkaido Regulatory Fisheries Research Laboratory,1983,48:53-77.
[15] Yatsu A,Watanabe T.Interannual variability in neon flying squid abundance and oceanographic conditions in the central North Pacific,1982-1992[J].Bulletion of the national research institute for far seas fisheries,1996,33:123-138.
[16] Yatsu A,Watanabe T,Mori J,et al.Interannual variability in stock abundance of the neon flying squid,Ommastrephes bartramii,in the North Pacific Ocean during 1979-1998:impact of driftnet fishing and oceanographic conditions[J].Fisheries oceanography,2000,9(2):163-170.
[17] 陈新军.北太平洋(160°E~170°E)大型柔鱼渔场的初步研究[J].上海水产大学学报,1999,8(3):197-201.
[18] 陈新军,曹杰,田思泉,等.表温和黑潮年间变化对西北太平洋柔鱼渔场分布的影响[J].大连水产学院学报,2010,25(2):119-126.
[19] 邵全琴,马巍巍,陈卓奇,等.西北太平洋黑潮路径变化与柔鱼CPUE的关系研究[J].海洋与湖沼,2005,36(2):111-122.
[20] 沈新强,樊伟,崔雪森.西北太平洋柔鱼渔场分布与水温关系的研究[J].海洋水产研究,2004,25(3):10-14.
[21] 陈新军,许柳雄.北太平洋150oE~165oE海域柔鱼渔场与表温及水温垂直结构的关系[J].海洋湖沼通报,2004,(2):36-44.
[22] USFWS.101 ESM Habitat as a Basis for Environmental Assessment[R].Washington,D.C:Division of Ecological Services U.S.Fish and Wildlife Service Department of the Interior,1980:4-80.
[23] Song L M,Zhou Y Q.Developing an integrated habitat index for bigeye tuna(Thunnus obesus)in the Indian Ocean based on longline fisheries data[J].Fisheries research,2010,105(2):63-74.
[24] Tian S Q,Chen X J,Chen Y,et al.Evaluating habitat suitability indices derived from CPUE and fishing effort data for Ommastrephes bartramii in the northwestern Pacific Ocean[J].Fisheries research,2009,95(2-3):181-188.
[25] 陈新军,刘必林,田思泉,等.利用基于表温因子的栖息地模型预测西北太平洋柔鱼(Ommastrephes bartramii)渔场[J].海洋与湖沼,2009,(6):707-713.
[26] 冯波,陈新军,许柳雄.应用栖息地指数对印度洋大眼金枪鱼分布模式的研究[J].水产学报,2007,31(6):805-812.
[27] 郭爱,陈新军.利用水温垂直结构研究中西太平洋鲣鱼栖息地指数[J].海洋渔业,2009,31(1):1-9.
[28] Chen X J,Tian S Q,Chen Y,et al.A modeling approach to identify optimal habitat and suitable fishing grounds for neon flying squid(Ommastrephes bartramii)in the Northwest Pacific O-cean[J].Fishery Bulletin,2010,108(1):1-14.
[29] 田思泉,陈新军,冯波,等.西北太平洋柔鱼资源丰度与栖息环境的关系及时空分布[J].上海海洋大学学报,2009,18(5):586-592.
[30] Andrade H A,Garcia C A E.Skipjack tuna fishery in relation to sea surface temperature off the southern Brazilian coast[J].Fisheries Oceanography,1999,8(4):245-254.
[31] Mohri M.Distribution of bigeye tuna in the Indian Ocean based on the Japanese tuna longline fisheries and survey information[D].Japan:National Fisheries University.1998.
[32] Hess G R,Bay J M.A regional assessment of windbreak habitat suitability[J].Environmental monitoring and assessment,2000,61(2):239-256.
[33] Lauver C L,Busby W H,Whistler J L.Testing a GIS model of habitat suitability for a declining grassland bird[J].Environmental Management,2002,30(1):88-97.
[34] 陈新军.关于西北太平洋的柔鱼渔场形成的海洋环境因子的分析[J].上海水产大学学报,1997,6(4):263-267.
[35] 陈峰,陈新军,刘必林,等.西北太平洋柔鱼渔场与水温垂直结构关系[J].上海海洋大学学报,2010,19(4):495-504.
[36] 陈峰,陈新军,钱卫国,等.水温变动对2009年西北太平洋柔鱼产量下降的影响[J].广东海洋大学学报,2010,30(1):65-71.
[37] Chen X J,Li G,Feng B,et al.Habitat suitability index of Chub mackerel(Scomber japonicus)from July to September in the East China Sea[J].Journal of Oceanography,2009,65(1):93-102.
[38] Zainuddin M,Kiyofuji H,Saitoh K,et al.Using multi-sensor satellite remote sensing and catch data to detect ocean hot spots for albacore(Thunnus alalunga)in the northwestern North Pacific[J].Deep Sea Research Part II:Topical Studies in Oceanography,2006,53:419-431.
