干旱对4种幼苗气体交换参数的影响
2012-01-04杨振意郭淑红
杨振意 , 薛 立 , 郭淑红 , 张 柔
(华南农业大学 林学院, 广东 广州 510642)
干旱对4种幼苗气体交换参数的影响
杨振意 , 薛 立 , 郭淑红 , 张 柔
(华南农业大学 林学院, 广东 广州 510642)
对蒲桃Syzygium jambos、南五味子Kadsura longipedunculata、海南红豆Ormosia pinnata和灰木莲Mangliet iaglauca幼苗在不同干旱强度下的叶片气体交换参数进行了研究。结果表明,随着干旱胁迫时间的延长,4种幼苗的净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)显著下降;蒲桃的胞间СO2浓度(Ci)显著降低,气孔限制值(Ls)下降后上升,南五味子和海南红豆的Ci都为下降后上升,Ls呈现上升后下降;灰木莲的Ci持续上升,而Ls持续下降。蒲桃的叶片水分利用率(WUE)上升,而其他幼苗则不断降低;主成分分析表明,幼苗耐干旱性顺序为蒲桃>南五味子>海南红豆>灰木莲。
干旱胁迫;幼苗;气体交换参数;主成分分析
光合作用是植物各项生命活动的基础,直接影响植物的生长和发育,其大小不仅与自身遗传特性有关, 而且受众多环境因子的影响[1]。干旱胁迫是树木逆境最普遍的形式[2],水是对光合作用有重要影响的环境因子之一,干旱条件下植物叶片的光合速率下降,生长受抑制。由于森林多生长在山地,少雨季节往往出现干旱胁迫,所以干旱胁迫下树木的光合作用一直是人们研究的热点。例如,Rouhi等[3]对干旱胁迫下的杏树Prunus armeniaca、Ogaya等[4]对冬青栎Quercus ilex和红景天Phillyrea latifolai、Ranjbarfardooei 等[5]对埃及黄连木Pistacia khinjuk和钝黄连木P. mutica光合速率等指标进行了研究;刘慧民等[6]研究了干旱胁迫的华北绣线菊和金山绣线菊光合能力,夏江宝等[7]报道了干旱胁迫的常春藤净光合速率日变化及其影响因子;罗明华[8]等研究了干旱胁迫对丹参叶片气体交换参数的影响。蒲桃Syzygium jambos叶密浓绿,树姿优美,花果期长,在华南地区广泛种植。南五味子Kadsura longipedunculata是具有较高的药用和食用价值的常绿或半常绿攀援木质藤本[9,10], 叶、花、果均可供观赏[10],具备优良的观赏特性和生态学特性[11]。海南红豆Ormosia pinnata是原产我国广东和海南岛以及越南等国的固氮乔木树种[12],主干通直, 萌芽更新力强, 耐土壤瘠薄, 冠形优雅,且抗风、防火、抗环境污染, 是优良的多用途树种[13]。灰木莲Mangliet iaglauca树形美观, 枝叶繁茂, 荫浓, 是优良的庭园及行道绿化树种。目前还未见干旱胁迫对这四种树种光合特性影响的报道。作者对这些内容进行研究,探讨其对不同水分状况的适应机制, 可以为山地造林时的树种选择提供参考。
1 材料与方法
1.1 材 料
2011年4月选择广东省林木种苗示范基地的1年生实生苗容器苗,放置华南农业大学林学院温室内进行研究。幼苗生长基本情况见表1。
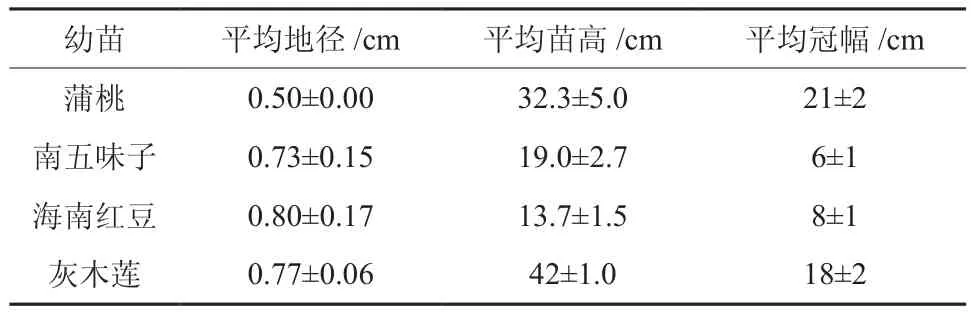
表1 4种幼苗的基本情况(平均值±标准差)Table 1 General characteristics of four seedlings(mean±SD)
1.2 方 法
采用不浇水自然干旱方式进行胁迫试验。在胁迫处理的0(对照)、2、4和6 d,进行光合气体交换参数和土壤容积含水量的测定。光合气体交换参数用美国 LІ-СOR 公司生产的 LІ-6400便捷式光合测定仪测定。测定参数主要包括:叶片净光合速率 (Pn,μmol•m-2·s-1)、气孔导度 (Gs,mmol•m-2·s-1)、 胞 间 СO2浓 度 (Ci,μmol•mol-1)和蒸腾速率 (Tr,mmol•m-2•s-1)。每株植物选择幼苗顶部开始的第3~8枚叶中的三枚成熟叶进行挂牌标记,测定时间为光合作用比较活跃的9:00~11:00,每叶片记录3个值,取平均值。测定时用红蓝光源叶室 , 光强 1000 µmol•m-2s-1, 温度为室温,空气流量为500 Lmol•s-1,СO2浓度为空气中СO2浓度。利用美国产MІ2X型土壤水分探头监测土壤容积含水量(RSVWС)的变化。计算水分利用效率RWUE和气孔限制值Ls计算公式分别见式1和式2:

式(2)中:Ca为大气СO2浓度。
为综合评价不同树种的抗旱性,采用SAS8.1软件系统对各项指标测定结果进行主成分分析,以确定4个树种的抗旱性[14]。
2 结果与分析
2.1 干旱胁迫中的土壤水分和幼苗形态变化
第0 、2、4和6d的土壤体积含水量分别为28%、14%、6%和3%(见图1)。在第2天时,蒲桃、南五味子、海南红豆和灰木莲形态无明显变化;第4 天时,蒲桃形态无明显变化,南五味子近根部分叶子成棕黄色,大部分叶子变皱并变软,海南红豆的小叶稍微闭合,灰木莲叶子开始萎焉变皱;第6天时,蒲桃叶片变化不大,叶片下垂,南五味子大部分叶子干枯蜷曲,海南红豆部分小叶基本闭合,有老叶脱落,灰木莲大部分叶片萎焉卷曲。
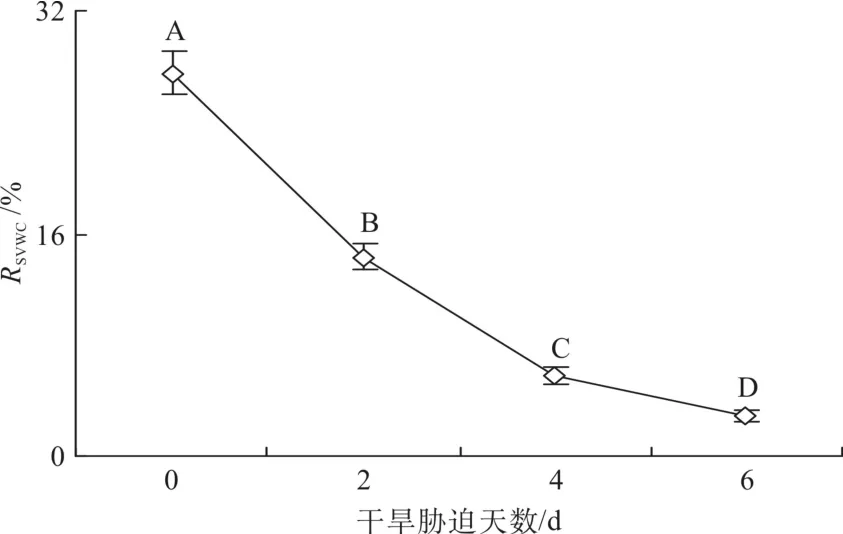
图1 干旱胁迫中的土壤体积含水量的变化Fig. 1 Changes of soil volumetric water content with time during drought stress
2.2 干旱胁迫中净光合速率的变化
由图2可知,第0天时蒲桃和南五味子的净光合速率(Pn)较高,海南红豆和灰木莲的Pn较低。随着干旱天数的增加,4种幼苗的Pn都不断降低,在第2d、4d和6d时与对照相比,蒲桃的Pn分别下降6%、85%和90%,南五味子分别下降7%、96%和104%,海南红豆分别下降6%、61%和90%,灰木莲分别下降29%、82%和90%。干旱胁迫过程中,蒲桃、南五味子和海南红豆的Pn在第4天开始下显著降低(P<0.001),灰木莲的Pn在第2天就开始显著降低(P<0.001)。在第6天时蒲桃的Pn相对最高,南五味子降为负值,海南红豆和灰木莲总降幅相差不大。

图2 干旱胁迫中净光合速率的变化Fig. 2 Changes of net photosynthetic rate during drought stress
2.3 干旱胁迫中气孔导度的变化
由图3可知,第0天时,蒲桃的气孔导度(Gs)最高,南五味子和灰木莲的Gs相近,海南红豆的Gs相对最低。随着干旱天数的增加,4种幼苗的Gs呈现与净光和速率相似的变化趋势,在第2天、4天和6天与对照相比,蒲桃的Gs分别下降8 %、88%和95%,南五味子的Gs分别下降25%、91%和92%,海南红豆的Gs分别下降11%、55%和84%,灰木莲的Gs分别下降32 %、75%和76%。胁迫过程中,蒲桃和海南红豆的Gs在第4天开始显著降低(P<0.001),南五味子和灰木莲在第2天就开始显著降低(P<0.001)。蒲桃和南五味子的Gs的总降幅相对较大,海南红豆次之,灰木莲最小。

图3 干旱胁迫中气孔导度的变化Fig. 3 Changes of stomatal conductance during drought stress
2.4 干旱胁迫中胞间СO2浓度的变化
由图4可知,在第0天时,蒲桃、南五味子和灰木莲的蒲桃的胞间СO2浓度(Ci)相差不大。随着干旱天数的增加,4种幼苗的Ci变化各异。胁迫过程中,蒲桃的Ci一直下降,在第2天、4天和6天 的Ci分别比对照相下降2%、4%和35%,南五味子的Ci分别为降低29%,升高36%和60%;海南红豆的Ci分别为降低11%,升高43%和56%;灰木莲的Ci分别为升高3%、18%和25%。南五味子和海南红豆Ci先降低后升高,灰木莲的Ci持续升高。

图4 干旱胁迫中胞间СO2浓度的变化Fig. 4 Changes of intercellular carbon dioxide concentration during drought stress
2.5 干旱胁迫中蒸腾速率的变化
由图5可知,第0天时,蒲桃的蒸腾速率(Tr)最高,南五味子和灰木莲的Tr相差近,海南红豆的Tr相对最低。随着干旱天数的增加,4种幼苗的Tr变化与气孔导度(Gs)的变化一致。与对照相比,蒲桃第2、4和6天的Tr分别下降5%、83%和93%,南五味子分别下降15%、88%和90%, 海南红豆分别下降10%、43%和78%,灰木莲分别下降27%、69%和70%。胁迫过程中,蒲桃和海南红豆的Tr在第4天显著降低(P<0.001),南五味子和灰木莲在第2天就开始显著降低(P<0.001)。蒲桃和南五味子的Tr总降幅相对最高,海南红豆与灰木莲的Tr总降幅相对较低。最终,蒲桃、南五味子和海南红豆的Tr较低,灰木莲的Tr最高。

图5 干旱胁迫中蒸腾速率的变化Fig. 5 Changes of transpiration rate during drought stress
2.6 干旱胁迫中气孔限制值的变化
由图6可知,第0天时,蒲桃、南五味子和灰木莲的气孔限制值(Ls)相差不大且相对较低,海南红豆的Ls相对较高。随着干旱天数的增加,4种幼苗的Ls表现出与胞间СO2浓度(Ci)相反的趋势。与对照相比,蒲桃第2、4和6d的Ls分别下降2%、上升12%和89%;南五味子分别升高41%,下降48%和94%;海南红豆分别升高5%,下降31%和46%;灰木莲分别下降2%、24%和38%。干旱过程中,蒲桃的Ls趋势为上升,南五味子和海南红豆Ls先下降后上升,灰木莲的Ls持续下降。
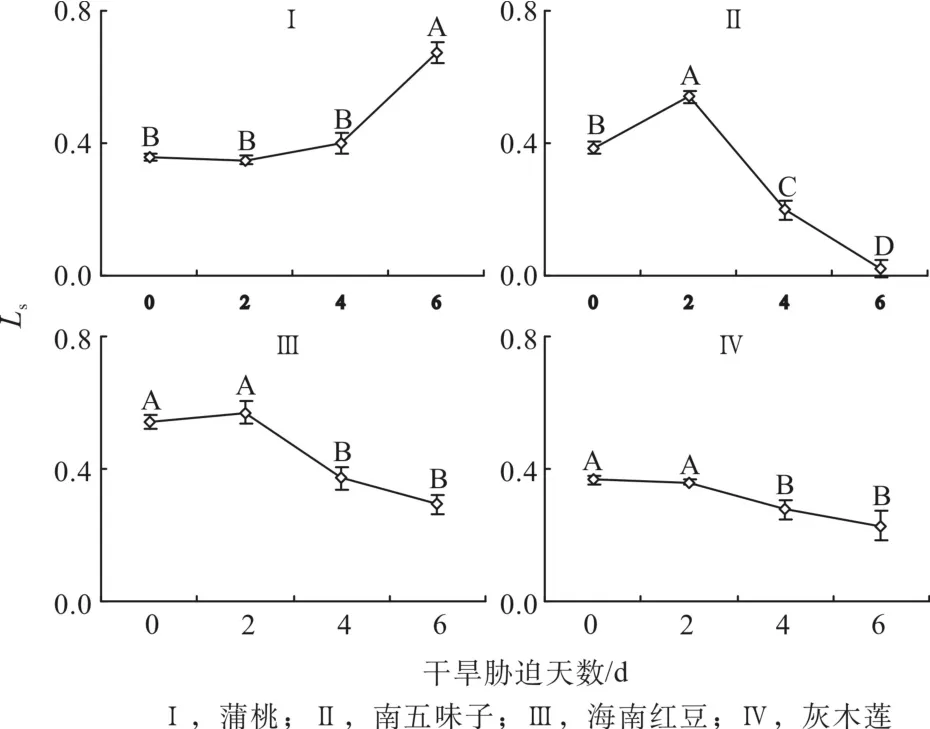
图6 干旱胁迫中气孔限制值的变化Fig. 6 Changes of stomatal limit during drought stress
2.7 干旱胁迫中叶片水分利用率的变化
由图7可知,第0天时,海南红豆的叶片水分利用率(RWUE)较高,南五味子、灰木莲和蒲桃相近。随着干旱天数的增加,4种幼苗的WUE变化不一。干旱胁迫第2天、4天和6 天与对照相比蒲桃的WUE相比分别为上升5%、3%和43%;南五味子的WUE分别下降7%、64%和100%,海南红豆的WUE分别降低13%、46%和50%,灰木莲的WUE分别降低7%、41%和53%。

图7 干旱胁迫中水叶片分利用率的变化Fig. 7 Changes of water use efficiency during drought stress
2.8 4种幼苗抗旱能力的综合评价
干旱胁迫下不同植物气体交换的差异体现了植物对干旱环境的适应能力[7],利用主成分对幼苗生理指标进行分析可以得出其抗旱能力[14]。由主成分的特征向量可计算出4种幼苗的主成分得分顺序为蒲桃、南五味子、海南红豆、灰木莲,蒲桃的主成分得分远远大于其他3种幼苗,表明其抗旱能力强。
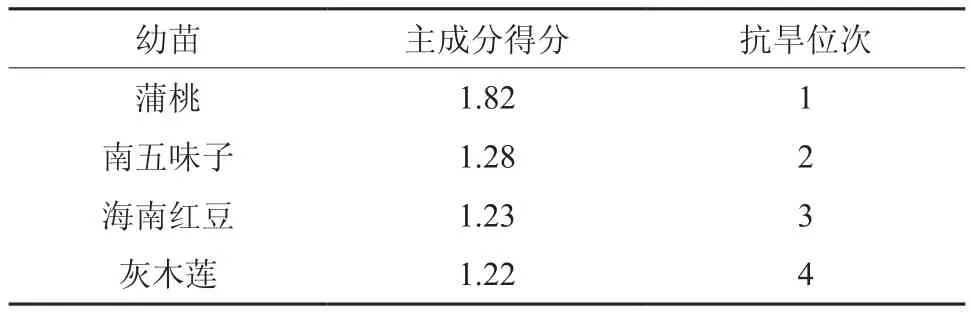
表1 干旱胁迫下4种幼苗的主成分得分Table 1 Score of principal components of 4 seedlings under drought stress
3 讨 论
本研究中,蒲桃、南五味子、海南红豆和灰木莲的净光合速率(Pn)、气孔导度(Gs)和蒸腾速率(Tr)随着干旱胁迫的加强都不断下降。植物受到干旱胁迫时,叶片水分散失和叶片水势下降,导致气孔开度减小,气孔阻力增加, 最后引起气孔关闭。气孔是光合作用时吸收空气中СO2的和蒸腾过程中水蒸气从体内排到体外的主要通道,干旱胁迫造成气孔阻力增加, СO2进入叶片受阻,同时光合产物输出变慢,叶片中淀粉水解加强,糖类积累,结果会引起光合速率下降[15,16]。植物干旱胁迫时通过关闭气孔而减少蒸腾失水,引起蒸腾速率下降,有利于推迟水分亏缺发展到有害或致死程度的时间,但是因为限制了СO2的进入而减弱光合作用。在干旱胁迫下,光合特性较早出现大幅度降低的幼苗,干旱适应性可能较差。本研究中,蒲桃的净光合速率(Pn)、气孔导度(Gs)和蒸腾速率(Tr)均在第4天显著下降,其他幼苗的这3种生理指标中,有的在第2天就显著降低,说明抗旱性强。
蒲桃的胞间СO2浓度(Ci)随着处理天数的增加持续降低,而气孔限制值(Ls)下降后升高,叶片形态未出现明显变化,说明气孔限制引起光合作用下降,但是叶片没有受损。南五味子和海南红豆的Ci均为下降后上升 ,Ls均呈现上升后下降;灰木莲的Ci为持续上升,Ls为持续下降。依据胞间СO2浓度(Ci)和气孔限制值(Ls)的变化方向[7]可以判断是气孔和非气孔限制因素[9]。气孔限制理论用胞间СO2浓度(Ci)和气孔限制值(Ls)的变化作为判别植物是否受气孔限制标准,若Ci降低和Ls增大, 则以气孔限制为主, 若Ci增大和Ls减小, 则以非气孔限制为主[7]。本研究中,限制蒲桃光合作用的主要是气孔限制,而海南红豆和南五味子在干旱2 d内 Ci降低和Ls增大,限制其光合作用的主要因素是气孔限制,因为轻度干旱胁迫造成根系导水能力下降而使气孔阻力增加,从而进一步限制СO2吸收,细胞间隙СO2浓度(Ci)下降和气孔限制值增加。此时光合能力的降低是短暂的可恢复性的, 对植物生理伤害较小[7]。海南红豆和南五味子干旱后2 d后,以及灰木莲在整个实验期间,Ci增大而Ls减小,说明Pn降低主要由气孔限制转向非气孔限制,幼苗木水分失调所引起的其他生理机能紊乱与反应机构的损伤使其物质代谢受阻[15],如叶绿体的电子传递速率降低且与光合磷酸化解偶联,影响同化力的形成,叶绿体变形,片层结构被破坏, 1, 5-二磷酸核酮糖(RuBP)的再生受到阻碍等[17]。海南红豆、南五味子和灰木莲的Pn大幅下降,叶肉细胞已经发生了不能逆转的变化,南五味子叶片在形态上已经表现出了巨大的变化,叶片干枯卷曲,光合作用基本停止,而海南红豆小叶叶片闭合,老叶开始脱落,灰木莲大部分叶片萎焉卷曲,这与前人对其他植物的研究结论基本一致[7,18-20]。
水分利用效率作为植物气体交换的一个重要指标,表征了植物在等量水分消耗情况下,固定СO2的能力,它不仅体现了植物自身光合能力大小,也能反映植物有效利用水分的能力[21],许多植物在干旱胁迫下通过改变自身的生理生态特性来达到高效用水的目的[21-23]。随着干旱天数的增加,蒲桃叶片的水分利用率持续增加,而南五味子、海南红豆和灰木莲的WUE持续降低,说明前者消耗同样数量的水分,可以固定较多的СO2,能在干旱胁迫下高效利用水分。
植物的抗旱性是一个复杂的综合性状,它受多种因素影响,不仅与植物的本身的遗传基因有关,而且还与植物的生长环境有关, 而各因素的重要性也不同,单一指标常常难以准确地评价某一树种的抗旱能力。冯慧芳等[14]研究4种苗木的抗旱生理指标和何跃君等[24]研究6种苗木的抗寒生理指标中,发现主成分分析的苗木抗逆能力与生理指标变化一致。本研究中,根据主成分分析的得分可知,4种幼苗的主成分分析抗旱得分顺序为蒲桃、南五味子、海南红豆、灰木莲,与其形态指标的变化一致,说明主成分分析适用于评价树木抗旱研究。
[1] 步秀芹,徐学选,杜 峰,等. 黄土丘陵区翻白草光合特性的研究[J]. 水土保持通报,2007, 27(6): 85-89.
[2] 李 燕, 薛 立, 吴 敏. 树木抗旱机理研究进展[J]. 生态学杂志,2007,26(11):1857-1866.
[3] ROUHІ V, SAMSON R, LEMEUR R, et al. Photosynthetic gas exchange characteristics in three different almond species during drought stress and subsequent recovery[J]. Environmental and Experimental Botany, 2007, 59: 117-129.
[5] RANJBARFORDOEІ A, SAMSON R, VAN DAMME P, et al. Effects of drought stress induced by polyethylene glycol on pigment content and photosynthetic gas exchange of Pistacia khinjuk and P. mutica[J]. Photosynthetica, 2000, 38(3): 443-447.
[6] 刘慧民,车艳双,车代弟,等. 干旱胁迫对华北绣线菊和金山绣线菊光合能力的影响[J]. 应用生态学报,2010, 21(8):2004-2009.
[7] 夏江宝,张光灿,许景伟,等. 干旱胁迫下常春藤净光合速率日变化及其影响因子分析[J]. 水土保持通报,2010, 30(3):78-82.
[8] 罗明华,胡进耀,吴庆贵,等. 干旱胁迫对丹参叶片气体交换和叶绿素荧光参数的影响[J]. 应用生态学报,2010, 21(3):619-623.
[9] 蓝盛芳. 中国南五味子属分类研究[J]. 华南农学院学报,1984, 5(2): 83-92.
[10] 邓白罗,谢碧霞,程 张. 中国南五味子属植物的种质资源及开发利用[J]. 中南林业科技大学学报,2008, 28(6): 90-94.
[11] 邓白罗,谢碧霞,刘 晖,等. 南五味子的地理分布与园林应用[J]. 中南林业科技大学学报,2009, 29(5): 184-186.
[12] 赵 平,曾小平,彭少麟,等. 海南红豆(Ormosia pinnata)夏季叶片气体交换、气孔导度和水分利用效率的日变[J]. 热带亚热带植物学报,2000, 8(1): 35-42.
[13] 粟 娟,孙 冰,王德祯. 海南红豆(Ormosia pinnata)的生物学特性和观赏价值[J]. 林业科学研究,1995, 8(6): 677-681.
[14] 冯慧芳,薛 立,任向荣,等. 4种阔叶苗木对PEG模拟干旱的生理响应[J]. 生态学报,2011,31(2): 371-382.
[15] YORDANOY І, TSONEY T, GOLTSEY V, et al. Іnteractive effect of water def i cit and high temperature on photosynthesis in sunf l ower and maize plants.1[J]. Photosynthetica, 1997, 33(3/4):391-402.
[16] 刘志媛,党选民,曹振木. 土壤水分对黄秋葵苗期生长及光合作用的影响[J]. 热带作物学报, 2003, 24(1): 70-73.
[17] FLEXAS J, MEDRANO H. Drought-inhibition of photosynthesis in С3 plant stomatal and non-stomatal limitations revisited[J].Annals of Botany, 2002, 89: 183-189.
[18] 窦新永,吴国江,黄红英,等. 麻疯树幼苗对干旱胁迫的响应[J]. 应用生态学报, 2008, 19(7): 1425-1430.
[19] 徐兴友,王子华,龙 茹,等. 干旱对6种野生花卉光合色素含量与气体交换的影响[J]. 经济林研究,2008, 26(4): 1-6.
[20] 杨玉珍,张云霞,彭方仁. 干旱胁迫对不同种源香椿苗木光合特性的影响[J]. 北京林业大学学报,2011, 33(1): 44-48.
[21] 闫海龙,张希明,许 浩,等. 塔里木沙漠公路防护林3种植物光合特性对干旱胁迫的响应[J]. 生态学报,2010, 30(10):2519-2528.
[22] GE Y, Lu Y J, LІAO J X. et al. Photosynthetic parameters of Mosla hangchowensis and Mosla dianthera as affected by soil moisture[J]. Photosynthetica, 2004, 42(3): 387-391.
[23] 包 卓,孟祥英,张晓松,等. 干旱胁迫对5 种园林绿化植物生理生化的影响[J]. 经济林研究,2010, 28(1): 46-50.
[24] 何跃君,薛 立,任向荣,等. 低温胁迫对6种幼苗生理特性的影响[J]. 生态学杂志,2008,27(4):524-531.
Effects of drought stress on leaf gas exchange parameters of four seedlings
YANG Zhen-yi, XUE Li, GUO Shu-hong, ZHANG Rou
(Сollege of Forestry, South Сhina Agricultural University, Guangzhou 510642, Guangdong, Сhina)
The effects of drought stress on leaf gas exchange of four seedlings ofSyzygium jambos,Kadsura longipedunculata,Ormosia pinnataandMangliet iaglaucawere studied. The results show that net photosynthetic rate (Pn), stomatal conductance (Gs)and transpiration rate (Tr) of the four seedlings continuously decreased with increasing drought time. Іntercellular carbon dioxide concentration(Ci) ofS. jambosdecreased, whereas its stomatal limits (Ls) fi rst decreased and then increased, whereas intercellular carbon dioxide concentration(Ci) decreased and then increased, theCiofK. longipedunculata,O. pinnatadecreased followed by an increase; theCiofM. iaglaucaincreased and itsLsdecreased during drought experiment period. Water use eff i ciency (WUE) ofS. jambosincreased and that of other seedlings decreased with increasing drought time. The drought resistance of the four seedlings was evaluated by using principal component analysis, and the order of drought resistance wasS. jambos>K. longipedunculata>O. pinnata>M. iaglauca.
drought stress; seedling; gas exchange parameter; analysis of principal component
S72;Q943
A
1673-923X (2012)05-0067-06
2011-11- 15
广东省林业局资助项目(F09054);广东省林业科技创新专项资金项目(2008KJСX008-01;2009KJСX011-01)
杨振意(1987-),男,硕士研究生,主要从事森林培育和树木生理学研究;E-mail:691514703@qq.com
薛 立(1958-),男,湖南桃江人, 教授,博士, 主要研究方向为森林生态和森林培育;E-mail: forxue@scau.edu.cn
[本文编校:欧阳钦]
