基于ISSR标记的南方红豆杉野生种群和迁地保护种群的遗传多样性和遗传结构分析
2011-12-31李乃伟贺善安束晓春
李乃伟,贺善安,束晓春,汪 庆,夏 冰,彭 峰
〔江苏省·中国科学院植物研究所(南京中山植物园),江苏 南京 210014〕
植物园以保存大量物种为特色,具有保存物种的“诺亚方舟”之美称。在木本植物方面,其迁地保护种群规模较小,往往只有少数几株,令人怀疑其在物种保护方面的实际作用与意义。从“最小有效种群”理论看,随着时间的推移,这些小种群都会消亡[1-2],所以,它们甚至被称为“活着的死植物”[3-4]。应该注意的是,这些理论和论断的依据都是自然或野生状态下小种群的动态规律,对于引种到植物园里的小种群,在人为条件影响下,其动态规律究竟如何则很少研究。李新华等[5]报道了在南京中山植物园内引种栽培40余年后南方红豆杉〔Taxus chinensis(Pilger) Rehd.var.mairei(Lemée et Lévl.)Cheng et L.K.Fu〕小种群的发展动态,由11株南方红豆杉组成的小种群,发展成1个具有400余植株的自然种群;在引种50余年后,该南方红豆杉小种群继续扩展到600株以上[6],南方红豆杉小种群的这一发展动态值得进一步研究。本文旨在对南方红豆杉小种群扩展后其遗传多样性的变化进行探讨,为了更广泛地研究南方红豆杉小种群在植物园内的动态,笔者等又在庐山植物园找到了类似的种群,更丰富了研究的内容。
种内遗传变异是物种适应性变异或进化的必要条件,对物种保护具有深远意义[7]。物种的进化潜力和抵御不良环境的能力既取决于种内遗传变异的大小,也有赖于遗传变异的种群结构[8-9]。迁地保护衍生种群和野生种群的遗传多样性研究具有重要的理论和实际意义,种群的遗传多样性越高或遗传变异越丰富,对环境变化的适应能力就越强,越容易扩展其分布范围和开拓新的环境[10]。
在南方红豆杉这一引种实例中,小种群衍生出的自然种群个体越来越多,衍生种群的遗传多样性丰富度是否发生变化以及怎样变化值得关注。为比较南方红豆杉迁地保护栽培种群及其衍生种群与野生种群在遗传多样性上的差异,作者选取与庐山地处同一个山脉——幕阜山脉的江西修水县黄沙镇和黄港镇的2个野生种群进行比较,并选用幕阜山脉之外的福建枫溪野生种群作为外种群,采用ISSR标记方法,比较了植物园次生林生境下南方红豆杉迁地保护栽培种群和衍生种群与野生种群的遗传多样性和遗传结构,以期为制定有效的濒危植物保护措施提供依据。
1 材料和方法
1.1 材料
于2007年7月至9月,在福建枫溪乡、江西黄沙镇和江西黄港镇分别选取3个南方红豆杉野生种群,在庐山植物园和南京中山植物园分别选取2个迁地保护栽培种群以及2个迁地保护衍生种群的实验材料,各种群的概况见表1。3个野生种群和庐山植物园迁地保护栽培种群各取19个单株(10雄9雌);南京中山植物园和庐山植物园的迁地保护衍生种群各取19个单株(雌雄未分化);南京中山植物园迁地保护栽培种群取8个单株(5雄3雌),共计122个单株。在每一单株上取当年生叶片,于硅胶中保存备用。
实验中使用的 d NTPs、10×PCR buffer及 Taq DNA聚合酶均购自宝生物工程(大连)有限公司; DNA marker购自南京天为生物有限公司;琼脂糖购自南京基威生物工程有限公司;所用引物由上海英骏生物技术有限公司合成。
1.2 方法
1.2.1 基因组总DNA的提取方法 取各单株的新鲜叶片,分别置于预冷的研钵中,加入液氮迅速磨成粉末,参照文献[11]的改良CTAB法提取基因组总DNA,并置于-20℃条件下保存、备用。
1.2.2 ISSR-PCR扩增条件及扩增结果的检测 扩增反应体系总体积20μL,含25 ng模板DNA、2.0μL 10×PCR buffer、2.0μL 25 mmol·L-1MgCl2、0.15 μL 2.5 mmol·L-1d NTPs、0.6μL 10μmol·L-1引物和0.2 U Taq DNA聚合酶。
用PE 9800 PCR仪(美国GeneAmp公司)进行扩增。ISSR-PCR扩增程序为:94℃预变性4 min;94℃变性30 s,58℃退火45 s,72℃延伸80 s,共38个循环反应;最后于72℃延伸5 min。

表1 供试的7个南方红豆杉种群概况Table 1 Status of seven populations of Taxus chinensis(Pilger)Rehd.var.mairei(Lemée et Lévl.)Cheng et L.K.Fu
ISSR-PCR扩增产物用质量体积分数1.2%的琼脂糖凝胶电泳检测,以2 000 bp DNA marker为参照,EB染色。电泳结果用FR-200A凝胶成像系统(上海复日科技有限公司)拍照。每个引物均重复扩增2次。
1.3 数据分析
对扩增产物稳定的8个引物的清晰条带进行统计分析,每个位点上有条带记为“1”,无条带记为“0”。利用POPGENE version 1.31软件[12]、参照文献[13-14]的方法分析南方红豆杉种群内和种群间各遗传多样性和遗传分化参数:多态性条带百分率(PPB)、Nei’s遗传多样性指数(h)、Shannon信息指数(I)和遗传分化系数(GST)。
2 结果和分析
2.1 ISSR扩增结果分析
7个南方红豆杉种群的ISSR扩增结果见表2。由表2可见,8个引物共扩增出条带73条,其中多态性条带62条;平均每个引物扩增出的条带数为9.1条,平均每个引物扩增出的多态性条带数为7.8条。

表2 用于南方红豆杉基因组总DNA ISSR标记分析的引物序列及扩增结果Table 2 Primer sequences used for ISSR marker analysis of total genom ic DNA from Taxus chinensis(Pilger)Rehd.var.mairei (Lemée et Lévl.)Cheng et L.K.Fu and am p lification results
2.2 基于ISSR标记的南方红豆杉种群的遗传多样性分析
采用ISSR标记方法分析了7个南方红豆杉种群的遗传多样性,相关参数见表3。7个南方红豆杉种群中,2个迁地保护衍生种群遗传多样性水平较高,其中南京中山植物园迁地保护衍生种群的多态性条带百分率(PPB)、Nei’s遗传多样性指数(h)和Shannon信息指数(I)值分别为64.38%、0.197 2和0.301 8;江西庐山植物园迁地保护衍生种群的PPB、h和I值分别为64.38%、0.189 5和0.291 1。江西黄港、江西黄沙和福建枫溪的3个野生种群的PPB、h和I值的总体水平居中,分别为57.53%~60.27%、0.148 3~0.188 3和0.233 8~0.283 1。相比之下,2个迁地保护栽培种群的遗传多样性水平较低,其中,南京中山植物园迁地保护栽培种群的PPB、h和I值分别为31.51%、0.107 8和0.161 3,江西庐山植物园迁地保护栽培种群的PPB、h和I值分别为56.16%、0.158 4和0.246 0。

表3 基于ISSR标记分析的7个南方红豆杉种群的遗传多样性分析Table 3 Analysis of genetic diversity of seven populations of Taxus chinensis(Pilger)Rehd.var.mairei(Lemée et Lévl.)Cheng et L.K.Fu based on ISSR marker analysis
2.3 南方红豆杉种群遗传多样性和遗传分化分析
将南方红豆杉的江西黄港种群(19个单株)、江西黄沙种群(19个单株)和福建枫溪种群(19个单株)3个野生种群合并作为1个野生种群(57个单株),江西庐山和江苏南京的迁地保护栽培种群合并作为1个迁地保护栽培种群(27个单株),江西庐山和江苏南京的迁地保护衍生种群合并作为1个迁地保护衍生种群(38个单株),采用POPGENE软件进行遗传多样性分析。
合并后的南方红豆杉野生种群、迁地保护栽培种群和迁地保护衍生种群的遗传多样性分析结果见表4。由表4可以看出,虽然3个野生种群以及2个迁地保护衍生种群各自的遗传多样性参数值,即多态性条带百分率(PPB)、Nei’s多样性指数(h)和Shannon信息指数(I)差异较大,但合并后的野生种群和迁地保护衍生种群的遗传多样性参数值比较接近,前者的PPB、h和I值分别为82.19%、0.205 8和0.325 9,后者的PPB、h和I值分别为78.08%、0.207 6和0.322 9;而迁地保护栽培种群的遗传多样性参数值相对较小,PPB、h和I值分别为60.27%、0.180 7和0.275 2。
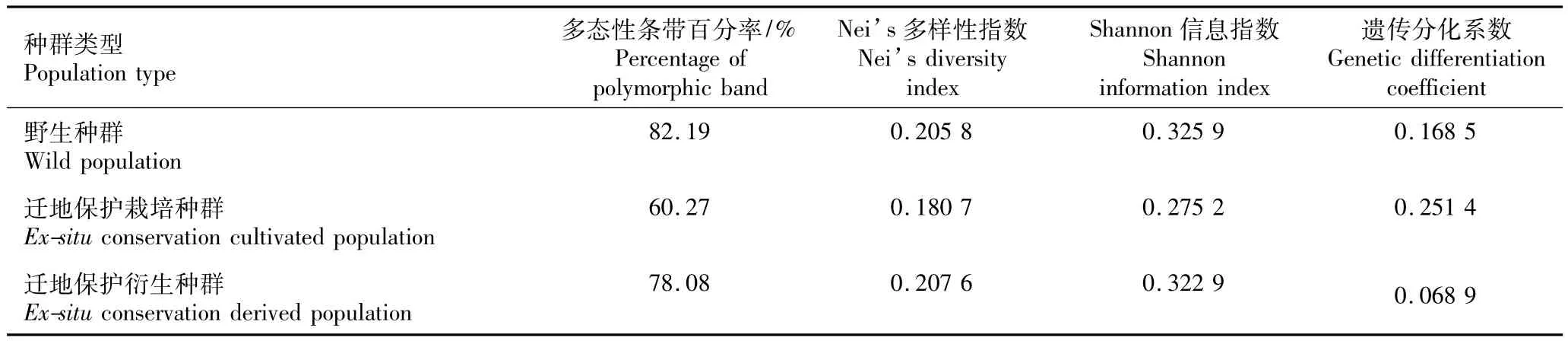
表4 南方红豆杉野生种群、迁地保护栽培种群和迁地保护衍生种群的遗传多样性和遗传分化参数Table 4 Genetic diversity and genetic differentiation parameters of w ild,ex-situ conservation cultivated and derived populations of Taxus chinensis(Pilger)Rehd.var.mairei(Lemée et Lévl.)Cheng et L.K.Fu
尽管合并后的南方红豆杉野生种群与迁地保护衍生种群的遗传多样性参数值相对接近,但两者的遗传分化系数(GST)差异较大,迁地保护衍生种群的GST为0.068 9,明显低于野生种群的GST(0.168 5);而迁地保护栽培种群的遗传分化系数最大(0.251 4),明显高于野生种群和迁地保护衍生种群。
3 讨 论
3.1 南方红豆杉3种种群的遗传多样性差异的比较
遗传多样性是生物多样性的核心,保护生物多样性最终是要保护其遗传多样性,因为一个物种的稳定性和进化潜力依赖其遗传多样性,而物种的经济和生态价值也依赖其特有的基因组成[15]。
本研究结果表明,根据Nei’s多样性指数(h),将南方红豆杉2个迁地保护衍生种群和3个野生种群分别在种群水平上进行分析,合并后的迁地保护衍生种群的遗传多样性与合并后的野生种群相近,单个迁地保护衍生种群的遗传多样性均较单个野生种群稍丰富,而合并后的或单个的迁地保护栽培种群的遗传多样性并不丰富。Shannon信息指数(I)同样说明了这种差异,合并后的迁地保护衍生种群、野生种群和迁地保护栽培种群的I值分别为0.322 9、0.325 9和0.275 2。对7个种群遗传多样性的比较结果表明,南京中山植物园迁地保护栽培种群的遗传多样性显著低于其他种群,其多态性条带百分率(PPB)、h和I值分别仅为31.51%、0.107 8和0.161 3。
南方红豆杉迁地保护衍生种群与迁地保护栽培种群之间产生的遗传多样性差异可能与其基因突变和遗传漂变有关,使得迁地保护衍生种群具有较高的遗传多样性。南京中山植物园的南方红豆杉迁地保护栽培种群的遗传多样性指标非常低,推测其原因有两方面:首先,样本数量较少,低于乔木遗传多样性分析所需的最低样本数15,如果能够达到庐山植物园迁地保护栽培种群的样本数量,这2个种群的遗传多样性指数可能会比较接近;而如果南京中山植物园和庐山植物园南方红豆杉的栽培数量再多一些,其遗传多样性指数可能也会接近野生种群。史全芬等[16]的研究结果表明,水杉(Metasequoia glyptostroboides Hu et W.C.Cheng)栽培种群的遗传多样性接近于野生种群,涵盖了野生种群近80%的遗传多样性。第二,南京中山植物园最初引种11株南方红豆杉,有3株在20世纪90年代死亡,但衍生种群中涵盖了这3株个体的遗传基因,这也可能会影响分析结果。
3.2 南方红豆杉迁地保护衍生种群与野生种群遗传结构的差异比较
植物种群的遗传结构反映了进化过程中的多种交互作用,如地理分布漂移、栖息地分裂、居群孤立、居群突变和基因漂变[17]。作者采用POPGENE软件统计分析ISSR标记的扩增结果,显示出南方红豆杉迁地保护衍生种群与野生种群不同的遗传结构。3个野生种群的遗传分化系数(0.168 5)接近于裸子植物的平均遗传分化系数(0.18)[18-19],而2个迁地保护衍生种群的遗传分化系数(0.068 9)则非常低。红豆杉科(Taxaceae)的北美红豆杉(Taxus brevifolia Nutt.)[20]、加拿大红豆杉(T.canadensis Marsh.)[21]和穗花杉〔Amentotaxus argotaenia(Hance)Pilg.〕[22]的野生种群的遗传分化系数都非常高。以上比较结果说明,孤立生长在2个植物园内的南方红豆杉迁地保护衍生种群与野生种群相比,前者种群间的遗传变异小于后者,进一步说明了植物园的“圈存”作用使迁地保护的植物种群更加孤立,增加了种群内部的变异,遗传漂变和基因突变的作用也更加明显。
3.3 南方红豆杉迁地保护的意义
遗传变异的保存是保护濒危物种的主要目的之一。种群内和种群间遗传变异的研究为濒危物种保存过程中形成合理的管理策略提供了重要信息[23]。本研究结果可以为南方红豆杉及红豆杉科其他植物的迁地保护提供参考。近年来,为了得到紫杉醇而对红豆杉科植物进行的乱砍乱伐,已经导致其濒危[24]。种群规模的缩小很容易导致这些物种由于随机的遗传漂变和同系繁殖而遭受遗传多态性丧失,保存这些濒危植物使其长期生存已经迫在眉睫。因此,南方红豆杉的迁地保护栽培种群、迁地保护衍生种群和野生种群的遗传结构的比较研究对该物种的保护有着重要的意义。
本研究中,尽管南方红豆杉迁地保护种群的遗传多样性较低,但所形成的迁地保护衍生种群的遗传多样性却出现了增加的趋势,说明植物园的迁地保护作用不可忽视,并不像以往有些推论,只能成为“活着的死植物”。所以,迁地保护可以成为一种有效的保护形式。然而,植物园作为濒危植物迁地保存的重要场所,应有自然林和人工林相结合的特殊群落结构,这种结构的存在能使11或29株母本植物繁殖后代形成一个具有丰富遗传多样性的天然种群。为使植物园保护的植物小种群能够长久繁衍,在建立一个新的植物园之初,就应考虑供游人参观的开放区与供植物种群繁衍扩张的自然区的结合。本文的研究结果对植物园的建园和管理策略提供了新的思考因素。
研究稀有物种或濒危物种种群的遗传变异在形成保护策略中有重要作用[25]。研究种群存活能力的生态学家Franklin认为,短期存活的有效种群大小不得低于50株[26];还有理论认为,采用迁地保护方式应该尽可能多的采集样本,如Lawrence认为迁地保护策略中至少收集172株植物[27],尤其应采集遗传多样性较高的种群内的样本。但是,本研究结果表明,含有少数个体的、遗传多样性较低的迁地保护小种群在保护物种上也具有相当重要的意义,其繁殖衍生出的新的自然小种群也有增加遗传多样性的可能。
目前,虽然南方红豆杉迁地保护衍生种群还有进一步扩张的趋势,但随着时间的推移,南方红豆杉个体之间是否会出现近亲繁殖导致的遗传退化,从而使衍生种群遗传多样性急剧降低进而影响其存活能力,还需要进一步研究。如果迁地保护衍生种群的后代种群不出现遗传变异的丧失,仍然保持遗传多样性的增长趋势,那么它们就能够适应环境的变化,容易存活下来进而扩展其分布范围。
[1]Barrett SCH,Kohn JR.Genetic and evolutionary consequences of small population size in plants:implications for conservation[M]∥Falk D A,Holsinger K E.Geneticsand Conservation of Rare Plants.New York:Oxford University Press,1991:3-30.
[2]Primack R,季维智.保护生物学基础[M].北京:中国林业出版社,2000:267.
[3]Janzen D H.Blurry catastrophes[J].Oikos,1986,47:l-2.
[4]贺善安,张佐双,顾 姻,等.植物园学[M].北京:中国农业出版社,2005:21.
[5]李新华,贺善安,盛 宁.红豆杉迁地保护中天然种群的形成[J].植物资源与环境,1999,8(1):38-41.
[6]贺善安,李新华,彭 峰,等.南方红豆杉迁地保护小种群动态的研究[J].植物资源与环境学报,2007,16(1):35-39.
[7]Schaal,B A,Leverich W J,Rogstad SH.A comparison ofmethods for assessing genetic variation in plant conservation biology[M]∥Falk D A,Holsinger K E.Genetics and Conservation of Rare Plants.New York:Oxford University Press,1991:123-134.
[8]Grant V.The Evolutionary Process:A Critical Study of Evolutionary Theory[M].2nd ed.New York:Columbia University Press,1991.
[9]Millar C I,Libby W J.Strategies for conserving clinal,ecotypic,and disjunct population diversity in widespread species[M]∥Falk D A,Holsinger K E.Genetics and Conservation of Rare Plants.New York:Oxford University Press,1991:149-170.
[10]Huenneke L F.Ecological implications of genetic variation in plant populations[M]∥Falk D A,Holsinger K E.Genetics and Conservation of Rare Plants.New York:Oxford University Press,1991: 31-34.
[11]田海燕,田新惠,李艳军,等.棉花DNA的提取及其SSR分子标记体系的建立[J].石河子大学学报:自然科学版,2007,25 (2):150-152.
[12]Yeh F C,Yang R C,Boyle T.POPGENE version 1.31:Microsoft Window-based Freeware for Population Genetic Analysis[CP/CD].Edmonton:University of Alberta,1999.
[13]Nei M.Analysis of gene diversity in subdivided populations[J].Proceedings of the National Academy of Sciences of the United States of America,1973,70(12):3321-3323.
[14]Lewontin R C.The apportionment of human diversity[J].Evolutionary Biology,1972,6:381-398.
[15]王洪新,胡志昂.植物的繁育系统、遗传结构和遗传多样性保护[J].生物多样性,1996,4(2):92-96.
[16]史全芬,杨 佳,李晓东,等.水杉栽培居群的遗传多样性研究[J].云南植物研究,2005,27(4):403-412.
[17]Schaal B A,Hayworth D A,Olsen K M,et al.Phylogeographic studies in plants:problems and prospects[J].Molecular Ecology,1998,7(4):465-474.
[18]Nybom H,Bartish I V.Effects of life history traits and sampling strategies on genetic diversity estimates obtained with RAPD markers in plants[J].Perspectives in Plant Ecology,Evolution and Systematics,2000,3(2):93-114.
[19]Nybom H.Comparison of different nuclear DNA markers for estimating intraspecific genetic diversity in plants[J].Molecular Ecology,2004,13(5):1143-1155.
[20]El-Kassaby Y A,Yanchuk A D.Genetic diversity,differentiation,and inbreeding in Pacific yew from British Columbia[J].Journal of Heredity,1994,85(2):112-117.
[21]Senneville S,Beaulieu J,DaoustG,etal.Evidence for low genetic diversity and metapopulation structure in Canada yew(Taxus canadensis):considerations for conservation[J].Canadian Journal of Forest Research,2001,31(1):110-116.
[22]Ge X J,Zhou X L,Li Z C,et al.Low genetic diversity and significant population structuring in the relict Amentotaxus argotaenia complex(Taxaceae)based on ISSR fingerprinting[J].Journal of Plant Research,2005,118(6):415-422.
[23]Avise JC,Hamrick J L.Conservation Genetics:Case Histories from Nature[M].New York:Chapman and Hall,1996.
[24]Milligan B G,Leebens-Mack J,Strand A E.Conservation genetics:beyond the maintenance of marker diversity[J].Molecular Ecology,1994,3(4):423-435.
[25]Francisco-Ortega J,Santos-Guerra A,Kim SC,etal.Plantgenetic diversity in the Canary Islands:a conservation perspective[J].American Journal of Botany,2000,87:909-919.
[26]Franklin IR.Evolutionary change in small populations[M]∥Soulé M E,Wilcox B A.Conservation Biology:An Evolutionary-Ecological Perspective.Sunderland:Sinauer Associates,Inc.,1980:135-149.
[27]Lawrence M J.A comprehensive collection and regeneration strategy for ex situ conservation[J].Genetic Resources and Crop Evolution,2002,49(2):199-209.
