芳樟不同无性系叶片光合色素含量及叶绿素荧光参数分析
2011-12-31林达定张国防于静波
林达定,张国防,于静波,冯 娟
(福建农林大学林学院,福建 福州 350002)
叶绿素含量是植物生理生化研究及生态调查中重要的测量参数之一,可用于表达植物生物量、检测植物健康程度及植物对环境的适应性[1]。叶绿素荧光动力学是将叶绿素荧光作为植物体内的天然探针,广泛用于植物光合作用机制、环境保护、作物增产潜力预测和植物逆境生理等研究领域[2-3]。植物生理特征与植物生长发育密切相关,不同生长发育阶段其生理特征出现相应的变化,而光是植物的能量源泉,对植物的生长发育和生理特征均有重要的影响[4]。
芳樟〔Cinnamomum camphora(L.)Presl〕又名香樟,是樟科(Lauraceae)樟属(Cinnamomum Trew)常绿乔木,是著名香料树种[5]。目前芳樟的研究与开发主要集中在形态特征、栽培技术、引种繁育、园林配置方式及化学成分[6]等方面,尚未见有关芳樟叶绿素荧光特性的报道。作者使用叶绿素荧光分析仪测定了18个芳樟无性系的叶绿素荧光参数并结合各无性系光合色素含量的测定结果,比较了芳樟不同无性系间的光能利用率、光合电子传递能力、光能转化效率和光化学反应效率的差异,以期对芳樟无性系间的光合生理状况进行综合比较,为芳樟优良无性系的选育提供理论依据。
1 材料和方法
1.1 材料
供试材料来源于福建省南平市郊教学林场(北纬26°58′~26°59′、东经118°12′~118°13′)5年生的芳樟无性系试验林,每个无性系种植5株。各无性系均采用条带状种植,株距和行距均为3 m。实验地气候属中亚热带气候,降水充沛,干、湿季分明,年平均气温19.3℃,立地条件适中。供试18个无性系分别为无性系79、114、116、187、195、209、242、264、283、BT1、BT2、BY2、MD1、PC1、PC2、PC4、PC5和WP1,均为根据芳樟叶含油率及芳樟醇含量选育而成。
1.2 研究方法
1.2.1 光合色素含量测定 于2010年7月初,在各无性系植株主干1/2高度的相同部位采集同一方向且长势相同的无病虫害叶片5~7片,快速洗净擦干并剪切成宽1~2 mm细条,混合均匀后称取0.5 g,共称取3份(视为3次重复);加入20 mL无水乙醇,密封置于黑暗处浸提1 d后将浸提液倒出,再加入15 mL无水乙醇,浸提至叶肉组织完全变白,合并2次浸提液,用无水乙醇定容至50 mL;以无水乙醇为空白,用UV-2401PC型紫外分光光度计(日本岛津公司)测定浸提液在波长470、649和665 nm处的光密度值,参照文献[7]的方法计算样品中光合色素含量。
1.2.2 叶绿素荧光参数测定 于2010年7月初,在各无性系植株主干1/2高度的相同部位选择同一方向且长势相同的无病虫害叶片3片进行标记,将叶片暗适应20 min,用叶绿素荧光分析仪(800MF/PSI便携式多谱动力学荧光成像系统,捷克PSI公司)对标记叶片进行离体测定,获取荧光参数:初始荧光(F0)、最大荧光(Fm)和可变荧光(Fv)=Fm-F0,并据此计算PSⅡ最大光化学效率(Fv/Fm)、PSⅡ潜在活性(Fv/F0)、PSⅡ电子传递情况 (Fm/F0)、光化学淬灭系数(Qp)、非光化学淬灭系数(NPQ)、PSⅡ实际光化学效率(Qy)和荧光下降比值(Rfd)等[8-9]。测定重复3次。
1.3 数据分析
采用DPS软件对所得数据进行聚类分析和方差分析,并采用LSD法进行多重比较。
2 结果和分析
2.1 芳樟无性系叶片光合色素含量的比较
叶绿素是绿色植物主要的光合色素,大部分叶绿素a和叶绿素b具有吸收及传递光能的作用,少数叶绿素a则具有在光反应中心负责将光能转化为化学能的作用;类胡萝卜素具有光能捕获和光破坏防御的功能,从而保护叶片合理吸收光能及协调光合作用[10-11]。芳樟不同无性系叶片中光合色素含量的比较结果见表1。由表1可以看出:芳樟不同无性系间的光合色素含量均存在差异,表明各无性系在吸收利用光能方面差异较大。各无性系间叶绿素b含量均小于叶绿素a含量,显示出叶绿素a在芳樟叶片吸收利用光能过程中起支配作用。其中无性系BT2叶片叶绿素a含量最高(2.26 mg·g-1),是无性系79 (0.60 mg·g-1)的3.8倍,说明无性系BT2吸收利用光能的能力较强;无性系MD1叶片中叶绿素a含量也较高,达到了2.17 mg·g-1。芳樟各无性系间类胡萝卜素含量的差异相对于叶绿素a和叶绿素b变化较小,类胡萝卜素含量多在0.2~0.3 mg·g-1,大部分无性系间的类胡萝卜素含量无显著差异,仅无性系242的类胡萝卜素含量显著高于无性系79、209、BY2、PC1、PC4和PC5,显示出不同无性系自我保护机制较一致。
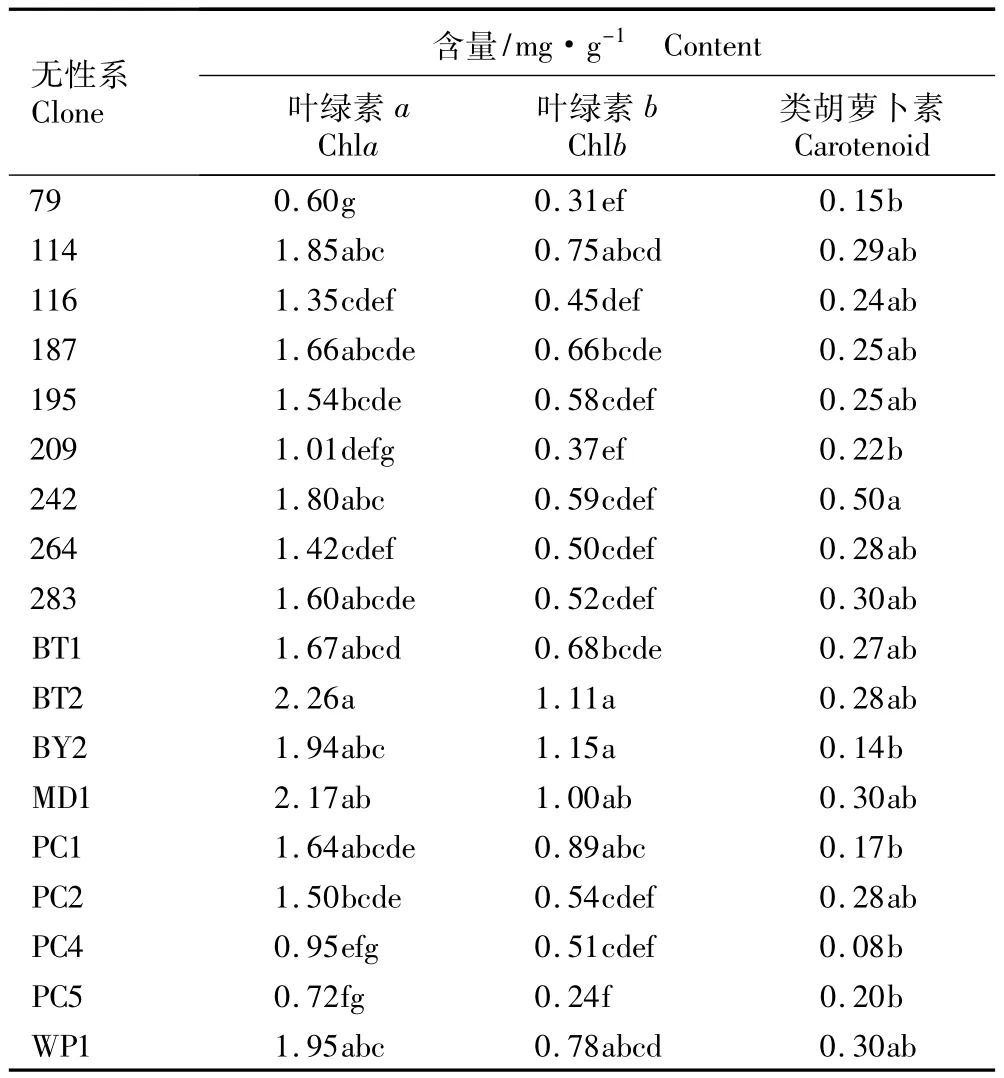
表1 芳樟不同无性系叶片中光合色素含量的比较1)Table 1 Comparison of photosynthetic pigment content in leaves of different clones of Cinnamomum camphora(L.)Presl1)
2.2 芳樟无性系叶片叶绿素荧光参数的差异性及相关性分析
2.2.1 叶绿素荧光参数的差异性分析 芳樟不同无性系叶片叶绿素荧光参数的多重比较结果见表2。由表2数据可见:不同无性系间叶片的初始荧光(F0)、PSⅡ最大光化学效率(Fv/Fm)、光化学淬灭系数(Qp)、非光化学淬灭系数(NPQ)、PSⅡ实际光化学效率(Qy)、PSⅡ潜在活性(Fv/F0)、PSⅡ电子传递情况(Fm/F0)存在一定差异,部分无性系间存在显著差异,但各无性系间的最大荧光(Fm)、可变荧光(Fv)和荧光下降比值(Rfd)的差异均不显著。
Fv/Fm值的变化代表光系统PSⅡ光化学效率的变化,常用来判断植物是否受到光抑制;在正常光照条件下,该值的波动范围在0.75~0.85,受到光抑制时低于0.75,该值越低证明植物受到光抑制的程度越高[12]。由表2可见:芳樟不同无性系的Fv/Fm值主要集中在0.80~0.85,显示出各无性系PSⅡ系统光能转换效率处于正常水平。无性系116、187和MD1的Fv/Fm值均高于其他无性系,说明这3个无性系光能转换效率较高。
Fm/F0可反映经过PSⅡ的电子传递情况;而Fv/F0可反映PSⅡ潜在的光化学活性,与有活性的PSⅡ反应中心数量成正比[13]。由表2可知:Fm/F0值与Fv/F0值的变化具有一致性。Fm/F0值越高,PSⅡ潜在的光化学活性越高,有活性的PSⅡ反应中心的数量也越多,能把所捕获的光能有效地转化为植物所需的化学能。无性系116、187、BT1和MD1的Fm/F0值和Fv/F0值均高于其他无性系,说明这4个无性系光能利用效果较好。
NPQ反映了植物热耗散能力的变化[14]。由表2可见:芳樟不同无性系间的热耗散能力差异显著,其中无性系BT2、PC5和PC1的NPQ值较高(分别为1.09、1.02和0.99),具有较强的热耗散能力,显示出这3个无性系具有较好的抗高温能力,而光能利用效果较差;无性系WP1和BY2的NPQ值相对较低(分别为0.19和0.31),热耗散能力较差,但能较充分利用光能。芳樟各无性系在非胁迫条件下均生长良好,因此各无性系具有较好的自我保护机制。
Qp反映了PSⅡ开放程度及原初电子受体QA的还原情况[14]。由表2可见:芳樟不同无性系的Qp值主要集中在0.77~0.87,利用光能效果总体较好,其中无性系PC5的Qp值最高(0.89),显示出其将光能转化为化学能的效果较好,可为植株生长提供较充足的能量保障。
Qy为实际量子产量,可作为植物叶片光合电子传递速率快慢的相对指标,表征植物目前的实际光合效率[14]。由表2可见:芳樟不同无性系的Qy值差异不明显,主要集中在0.47~0.55,与Fv/Fm值相差约0.3,说明各无性系把叶片所捕获的光能转化为植株所需的化学能的效率较低,其中无性系116、209和114的Qy值分别为0.55、0.54和0.54,这3个无性系的PSⅡ实际光化学效率相对较高。
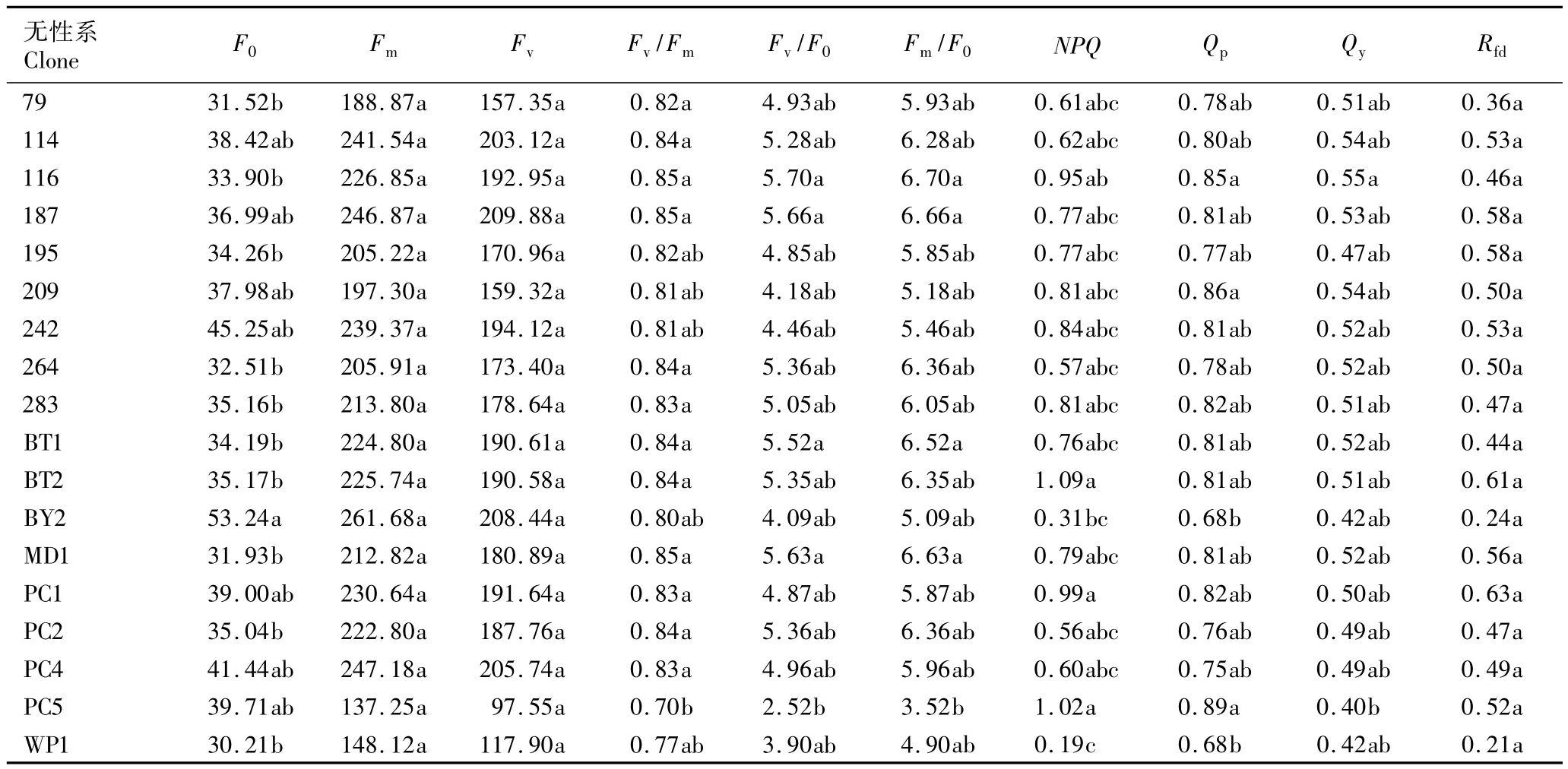
表2 芳樟不同无性系叶片叶绿素荧光参数的比较1)Table 2 Com parison of chlorophyll fluorescence parameter in leaves of different clones of Cinnamomum camphora(L.)Presl1)
2.2.2 叶绿素荧光参数的相关性分析 芳樟不同无性系叶片叶绿素荧光参数的相关性分析结果见表3。由表3可见:叶片的初始荧光(F0)仅与最大荧光(Fm)显著正相关,与其他参数的相关性不显著,说明初始荧光产量值将影响最大荧光产量;光化学淬灭系数(Qp)与非光化学淬灭系数(NPQ)和荧光下降比值(Rfd)存在极显著正相关,与 PSⅡ实际光化学效率(Qy)呈不显著的正相关,与其他参数呈负相关,说明芳樟不同无性系PSⅡ开放程度及原初电子受体QA的还原情况将影响植物热耗散能力;Qy与可变荧光(Fv)、PSⅡ最大光化学效率(Fv/Fm)、PSⅡ潜在活性(Fv/F0)、PSⅡ电子传递情况(Fm/F0)极显著正相关,显示出芳樟不同无性系叶片的实际光合效果主要由可变荧光值的多少决定;Rfd与Qy、Qp、NPQ存在显著或极显著正相关,显示出芳樟不同无性系叶片潜在光合作用量子转化效率将影响植物实际光合效率、植物热耗散能力、PSⅡ开放程度及原初电子受体QA的还原情况。因此,叶片潜在光合作用量子转化效率可以用于研究芳樟叶片实际光能利用情况。
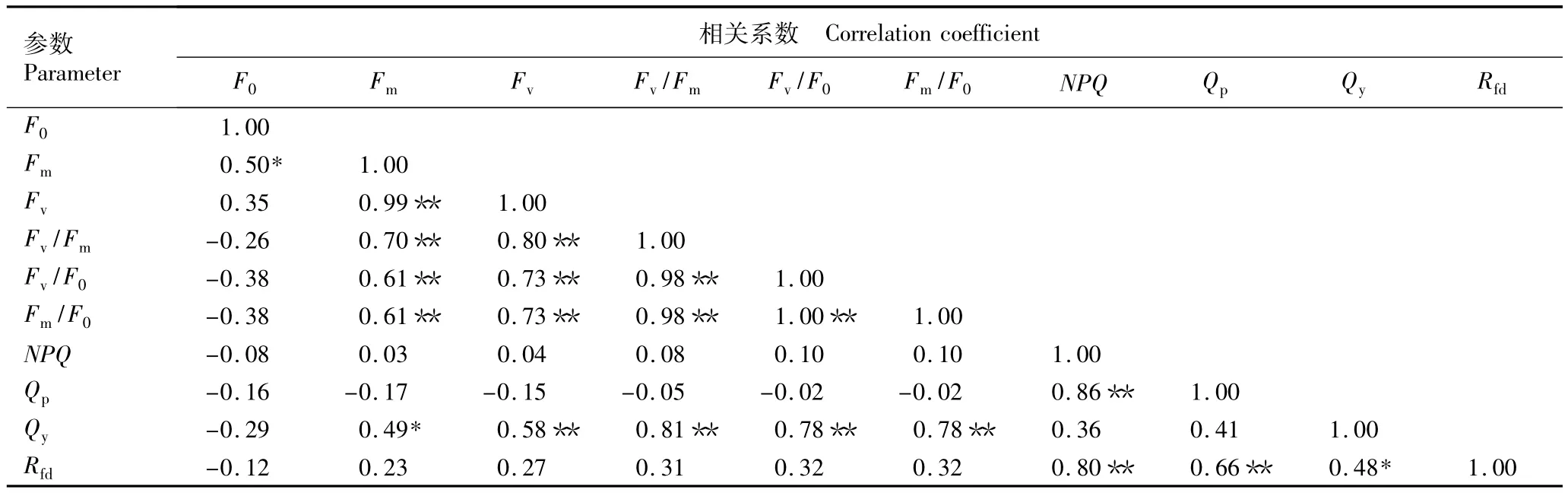
表3 芳樟不同无性系叶片叶绿素荧光参数的相关性分析1)Table 3 Correlation analysis of chlorophyll fluorescence parameter in leaves of different clones of Cinnamomum camphora(L.)Presl1)
另外,表3中Fv/Fm与Fv/F0、Fm/F0的相关系数均达0.98,说明芳樟不同无性系叶片PSⅡ潜在的光化学活性及电子传递情况将影响PSⅡ光能转换效率;Fm/F0与Fv/F0相关系数达到1.00,说明PSⅡ的电子传递情况将决定PSⅡ潜在的光化学活性。
2.3 芳樟无性系的聚类分析
根据芳樟不同无性系叶片的光合色素含量及叶绿素荧光参数测定结果,采用欧氏距离及最短距离聚类法,获得了18个芳樟无性系的聚类图(图1)。18个无性系可明显划分为2类:第1类共包含16个无性系;第2类仅包含无性系PC5和WP1。第1类又可分为2个亚类:第1亚类仅包含1个无性系BY2;第2亚类包含195、264、MD1、283、PC4、187、114、PC1、PC2、BT2、BT1、116、242、209和79等15个无性系,其中无性系BT1与BT2距离最短,光合生理特征最接近;无性系195与264、MD1与283、PC4与187、209与79也具有相近的光合生理特征。聚类结果说明:供试的大多数芳樟无性系叶片光合生理特征总体较相近。
3 讨论和结论
叶绿素是植物主要的光合色素,在光合作用中以电子传递及共振的方式参与能量的传递反应,其含量变化与植物生长发育有关[15]。光合机构中叶绿素吸收的光能主要用于推动光合作用,也往往有一部分在形成同化力之前以热的形式耗散和以荧光的形式重新发射出来[16-17]。芳樟不同无性系叶片叶绿素a和叶绿素b、类胡萝卜素含量及叶绿素荧光参数均存在一定差异,甚至部分无性系间存在显著差异,说明芳樟不同无性系对光能利用存在差异,且在不同无性系叶片吸收利用光能的过程中叶绿素a起决定性作用。根据非光化学淬灭系数(NPQ)和光合色素含量较高的特点,判断无性系BT2能够较好地利用光能,同时也具有较好的热耗散能力。无性系BT2在充分利用光能的同时,能将剩余的光能以热的形式耗散,从而保护芳樟的光合机构,具有较好的自我保护机制,这与贺立红等[18]对银杏(Ginkgo biloba L.)不同品种叶绿素荧光参数的研究结果类似。何炎红等[19]认为:非光化学淬灭耗散变化与光合速率变化具有正相关性。可见无性系BT2具有良好的光合生理性能,可为植株生长提供较充裕的化学能,同时可保护光合机构免受损坏。因此,可以通过对叶绿素荧光参数的观测来探究光合作用和热耗散的状况。
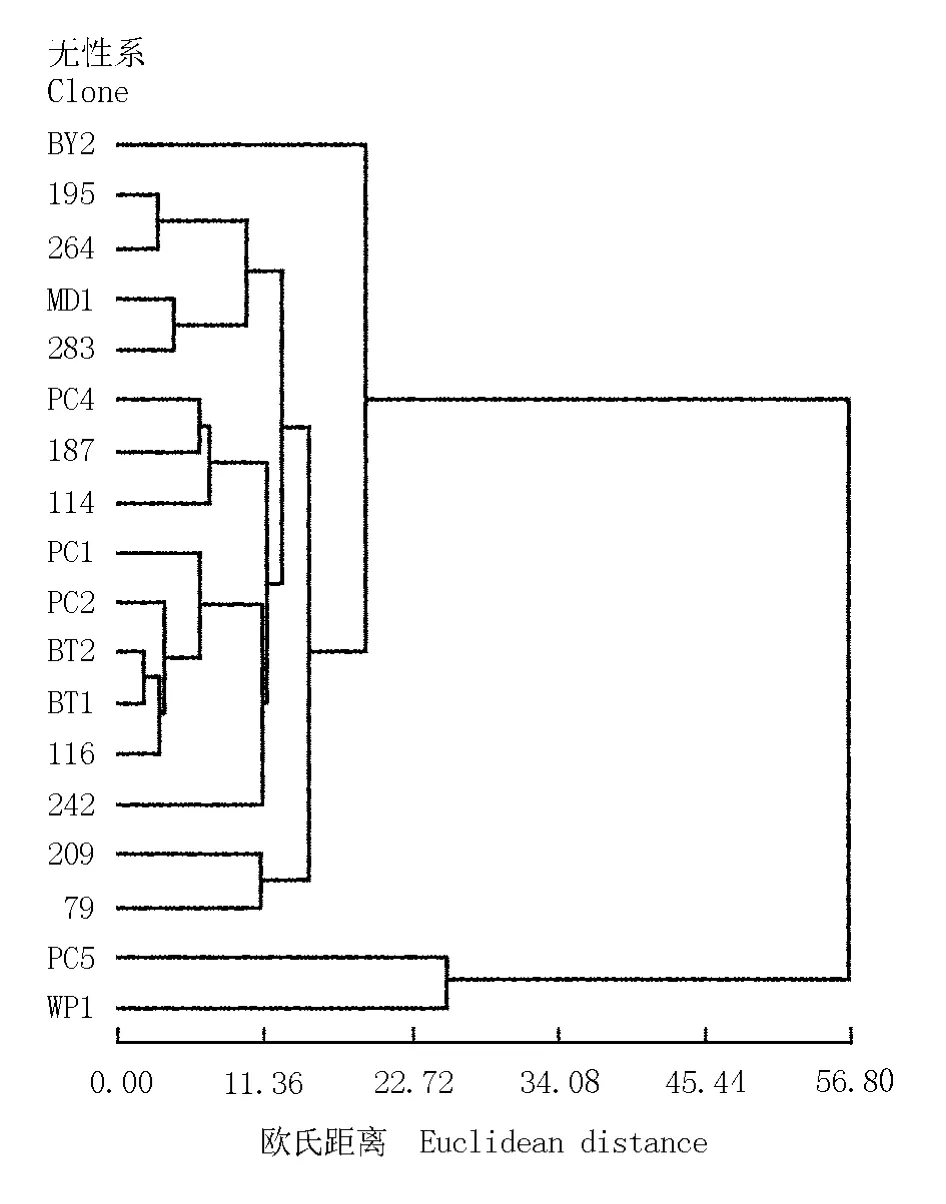
图1 基于光合色素含量和叶绿素荧光参数测定结果的18个芳樟无性系聚类分析图Fig.1 Cluster diagram of eighteen clones of Cinnamomum camphora (L.)Presl based on determ ination results of photosynthetic pigment content and chlorophyll fluorescence parameter
Fv/Fm和Qy是叶绿素荧光动力学分析的2个重要参数,Fv/Fm表征光化学反应状况及PSⅡ反应中心的最大光能转换效率,Qy则反映植物目前的实际光合效率。研究结果表明:芳樟不同无性系叶片PSⅡ原初光化学反应的最大量子光化学效率为0.75~0.85,表明其光合机构光能转换的效果较好[20],具有较大的光化学耗散能量的潜在能力。根据Qy值可知:芳樟不同无性系叶片实际光能利用效果差别较大,且总体上能把所捕获的光能转化为植株所需化学能的效率较低。吕芳德等[9]认为:主要由于PSⅡ的电子传递情况差异影响PSⅡ潜在的光化学活性,从而影响叶片吸收利用光能的比例。
Rfd能够表征叶片潜在光合作用量子转化效率。相关性分析结果表明:芳樟不同无性系叶片潜在光合作用量子转化效率将影响叶片实际光合效率、热耗散能力、PSⅡ开放程度及原初电子受体QA的还原情况等4个方面。余叔文等[21]认为:光合碳同化的量子效率与PSⅡ的光化学效率之间有很好的直线关系,光合碳同化的量子效率将影响光化学效率的高低。因此,叶片潜在光合作用量子转化效率可以用于研究芳樟实际光能利用情况,对深入研究芳樟光合生理变化机制具有重要意义。
[1]张友胜,张苏峻,李镇魁.植物叶绿素特征及其在森林生态学研究中的应用[J].安徽农业科学,2008,36(3):1014-1017.
[2]徐德聪,吕芳德,潘晓杰.叶绿素荧光分析技术在果树研究中的应用[J].经济林研究,2003,21(3):88-91.
[3]彭方仁,朱振贤,谭鹏鹏,等.NaCl胁迫对5个树种幼苗叶片叶绿素荧光参数的影响[J].植物资源与环境学报,2010,19(3): 42-47.
[4]潘瑞炽.植物生理学[M].4版.北京:高等教育出版社,2001: 61-63.
[5]中国科学院中国植物志编辑委员会.中国植物志:第三十一卷[M].北京:科学出版社,1982:182-184.
[6]张国防,陈存及.福建樟树叶油的化学成分及其含量分析[J].植物资源与环境学报,2006,15(4):69-70.
[7]李合生.植物生理生化实验原理和技术[M].2版.北京:中国农业出版社,2001:45.
[8]罗 俊,张木清,林彦铨,等.甘蔗苗期叶绿素荧光参数与抗旱性关系研究[J].中国农业科学,2004,37(11):1718-1721.
[9]吕芳德,徐德聪,侯红波,等.5种红山茶叶绿素荧光特性的比较研究[J].经济林研究,2003,21(4):4-7.
[10]孙小玲,许岳飞,马鲁沂,等.植株叶片的光合色素构成对遮阴的响应[J].植物生态学报,2010,34(8):989-999.
[11]吕福梅,沈 向,王东生,等.紫叶矮樱叶片色素性质及其光合特性研究[J].中国农学通报,2005,21(2):225-228.
[12]何炎红,郭连生,田有亮.7种针阔叶树种不同光照强度下叶绿素荧光猝灭特征[J].林业科学,2006,42(2):27-31.
[13]杜亮亮,金爱武,胡元斌,等.5种箬竹属竹种叶绿素荧光特性的比较[J].世界竹藤通讯,2009,7(2):17-21.
[14]冯建灿,胡秀丽,毛训甲.叶绿素荧光动力学在研究植物逆境生理中的应用[J].经济林研究,2002,20(4):14-18,30.
[15]李保印,周秀梅,王西波,等.不同彩叶植物叶片中叶绿体色素含量研究[J].河南农业大学学报,2004,38(3):285-288.
[16]黄华宏,陈奋学,童再康,等.矮生杉木光合特性及叶绿素荧光参数研究[J].北京林业大学学报,2009,31(2):69-73.
[17]冯立国,俞 菊,陶 俊,等.高温胁迫对一品红光合作用与叶绿素荧光的影响[J].扬州大学学报:农业与生命科学版,2009,30(3):71-74.
[18]贺立红,贺立静,梁 红.银杏不同品种叶绿素荧光参数的比较[J].华南农业大学学报,2006,27(4):43-46.
[19]何炎红,田有亮,郭连生.几种针阔叶树种不同光照强度下叶绿素荧光特性的研究[J].生态学杂志,2005,24(5):467-472.
[20]许大全,张玉忠,张荣铣.植物光合作用的光抑制[J].植物生理学通讯,1992,28(4):237-243.
[21]余叔文,汤章城.植物生理与分子生物学[M].2版.北京:科学出版社,1998:262-267.
