皖西大别山多枝尖山区主要乔木树种种间联结特征
2011-12-31王志高吴甘霖段仁燕张中信沈三保
王志高,吴甘霖,①,段仁燕,张中信,沈三保
探讨环境因素对植物分布和群落构建的影响是植物生态学研究的基本内容之一[1]。群落生境的差异影响物种分布、导致不同物种在空间分布上产生相互关联性,也即种间联结 (species association)[2-4]。种间联结分析通常以物种是否存在二元数据为依据[5-15],研究种间相互吸引或排斥的性质[2-3]、探讨植物对环境的功能群适应能力[16-17],有助于更好地了解群落中各个物种之间的相互作用以及群落的组成、动态和物种多样性的维持机制等[3-4]。
多枝尖山区是皖西大别山腹地主峰分水岭的主段,最高峰海拔 1 721.5m。该区域的地带性植被为常绿 -落叶阔叶混交林,植物区系起源古老,属华中、华西、华北、华南和华东植物区系的交会地带,物种特有现象明显,是大别山植物区系物种最丰富的地区之一[17-18]。该区域的森林植被在 20世纪 70年代曾遭到大规模砍伐,目前是鹞落坪国家级自然保护区的核心区之一,区内次生植被恢复状况良好。有关该区域植物区系和植被特征的研究已有较多的报道[19-22],但缺乏对群落结构及其动态变化的进一步研究。
作者采用X2检验、联结系数和共同出现百分率 3个指标对多枝尖山区主要乔木树种的种间联结特征进行了分析,并据此探讨了该区域植物群落中主要乔木树种之间的相互关系,以期为进一步研究该区域植物群落特征及其动态变化奠定基础。
1 研究区概况和研究方法
1.1 研究区概况
大别山多枝尖山区位于安徽省岳西县境内,地理坐标为东经 116°03′~116°09′、北纬 31°00′~31°11′,总面积约 210 km2。区域内的自然条件和植被概况参见文献[18-21]所述。
1.2 研究方法
1.2.1 样方设置和数据调查方法 采用样方法进行群落调查。共设置 25个面积为 20m×20m的典型乔木样方,每个乔木样方中分别随机设置5个面积为2m×2m的灌木样方和 5个面积为 1m×1m的草本样方。乔木层采用逐株调查方法进行调查,记录每木的种名、胸径和坐标;灌木层和草本层则分别记录各种类的种名、株数、高度和盖度,同时还记录了各个乔木样方的坡度、坡向和海拔等环境数据。
1.2.2 种间联结分析的指标及计算方法 由于该区域很多植物种类的出现频率较低,而出现频率较小的种类种间联结性也较小,因此,仅对出现频率大于5%的 30个乔木种类的种间联结性进行分析。
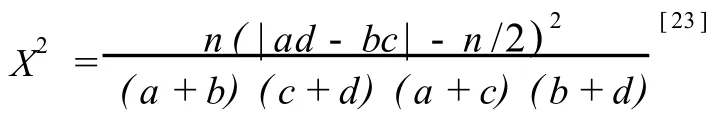
联结系数 (association coefficient,AC)用于进一步检验X2检验的结果及说明种间联结程度,计算公式[4-13]如下:ad≥bc时,AC=(ad-bc)/〔(a+b)(b+d)〕;bc>ad且d≥a时,AC=(ad-bc)/〔(a+b) (a+c)〕;bc>ad且d<a时,AC=(ad-bc)/〔(b+d)(d+c)〕。AC值域为[-1,1]。AC值越趋近于1,表明 2个物种间的正联结性越强,即 2个物种共同出现或共同不出现的可能性越大;AC值越趋近于 -1,表明 2个物种间的负联结性越强,即 2个物种单独出现的可能性越大;AC值为 0,则表示 2个物种完全独立,彼此之间没有任何联系。AC值只能反映种间联结性的强弱,无法判断其显著水平。
共同出现百分率(percentage of co-occurrence,PC)用于测度物种间的正联结程度,计算公式[4-13]为:PC=a/(a+b+c)。PC值域为[0,1]。PC值越趋近于 1,表明种对间的正联结性越强,即 2个物种共同出现或共同不出现的可能性越大,间接反映出 2个物种的生态习性及对环境的需求越一致。与AC值相似,PC值也无法反映种间联结性的显著水平。
2 结果和分析
根据皖西大别山多枝尖山区主要乔木树种的X2检验结果绘制的半矩阵图见图 1;根据联结系数(AC)分析结果绘制的半矩阵图见图 2;根据共同出现百分率(PC)分析结果绘制的半矩阵图见图 3。
2.1 X2检验分析
图 1结果表明:皖西大别山多枝尖山区的 30种主要乔木树种组成的 435个种对中有 23个种对的种间联结性达到显著 (P<0.05)或极显著 (P<0.01)水平,其他种对间的种间联结性均不显著,反映出该植物群落中乔木种对间的种间联结较松散。
正联结的种对有 15个,其中,华东椴 (Tilia japonicaSimonk.)与安徽槭 (AceranhweienseFang et Fang f.)和省沽油 (StaphyleabumaldaDC.)、黄山栎(QuercusstewardiiRehd.)与楤木 (AraliachinensisL.)、短柄枹(QuercusglanduliferaB lum e)与华山矾〔Symplocoschinensis(Lour.)D ruce〕和山樱花〔Cerasus serrulata(L ind l.)G.Don ex London〕、鹅耳枥(CarpinusturczaninowiiHance)与漆树〔Toxicodendron vernicifluum(Stokes)F.A.Bark ley〕和四照花〔Dendrobenthamiajaponica(DC.)Fang var.chinensis(Osborn)Fang〕、华千金榆 (CarpinuscordataB lum e var.chinensisFranch.)与 绒 毛 石 楠(Photinia schneiderianaRehd.etW ils.)、华山矾与水榆花楸〔Sorbusalnifolia(Sieb.et Zucc.)K.Koch〕、化香(PlatycaryastrobilaceaSieb.et Zucc.)与山樱花之间的联结性达到显著水平 (P<0.05);安徽槭与黄山栎和省沽油、华山松 (PinusarmandiiFranch.)与雷公鹅耳枥 (CarpinusvimineaL ind l.)、黄山栎与省沽油、金钱松〔Pseudolarixamabilis(J.Nelson)Rehd.〕与杉木〔Cunninghamialanceolata(Lam b.)Hook.〕5个种对的种间联结性达到极显著水平(P<0.01)。

图 1 皖西大别山多枝尖山区主要乔木树种种间联结分析的 X2半矩阵图F ig.1 X2-sem im a tr ice d iagram of in ter-spec ific assoc ia tion ana lysis of dom inan tarbor species in Duozh ijianm oun ta inous reg ion of Dab ieshan M oun ta in inW est Anhu i
负联结的种对有 8个,其中,安徽槭与黄山松(PinustaiwanensisHayata)、鹅耳枥与香槐 (Cladrastis wilsoniiTakeda)、黄山栎与茅栗 (CastaneaseguiniiDode)和山樱花、黄山松与省沽油、杉木与四照花之间的联结性达到显著水平(P<0.05);茅栗与安徽槭和省沽油之间的联结性达到极显著水平(P<0.01)。
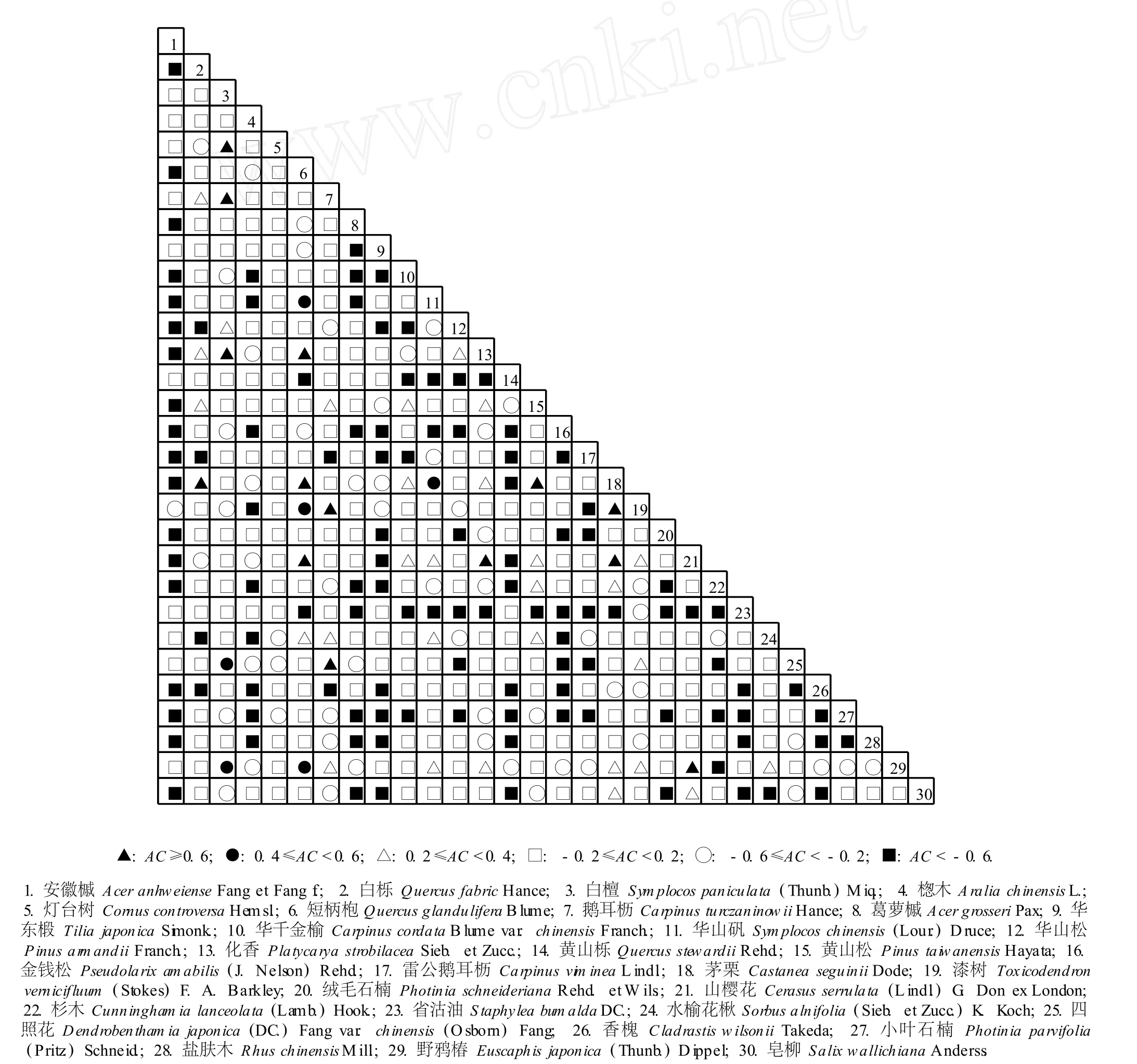
图2 皖西大别山多枝尖山区主要乔木树种种间联结分析的联结系数(AC)半矩阵图F ig.2 A ssoc ia tion coeffic ien t(AC)-sem im a tr ice d iagram of in ter-spec ific assoc ia tion ana lysis of dom inan tarbor spec ies in Duozh ijian m oun ta inous reg ion of Dab ieshan M oun ta in in W est Anhu i
2.2 联结系数(AC)分析
图 2结果表明:皖西大别山多枝尖山区植物群落中联结系数(AC)值在 0.6以上 (AC≥0.6)的乔木种对共有 14个,占主要乔木种对总数的 3.2%,这 14个种对分别为白栎 (QuercusfabricHance)与茅栗,白檀〔Symplocospaniculata(Thunb.)M iq.〕与灯台树(CornuscontroversaHem sl.)、鹅耳枥和化香,短柄枹与化香、茅栗和山樱花,鹅耳枥与漆树和四照花,化香与山樱花,黄山松与茅栗,茅栗与漆树和山樱花,山樱花与野鸦椿〔Euscaphisjaponica(Thunb.)D ippel〕;AC值在 0.4~0.6(0.4≤AC<0.6)的种对有 6个,占主要乔木种对总数的 1.4%,这 6个种对分别为白檀与四照花和野鸦椿,短柄枹与华山矾、漆树和野鸦椿,华山矾与茅栗;AC值在 0.2~0.4(0.2≤AC<0.4)的种对有29个,占主要乔木种对总数的 6.7%;AC值在 -0.2~ 0.2(-0.2≤AC<0.2)的种对有 213个,占主要乔木种对总数的 49.0%;AC值在-0.6~-0.2(-0.6≤AC<-0.2)的种对有60个,占主要乔木种对总数的13.8%;AC值低于 -0.6(AC<-0.6)的种对有 113个,占主要乔木种对总数的 26.0%。
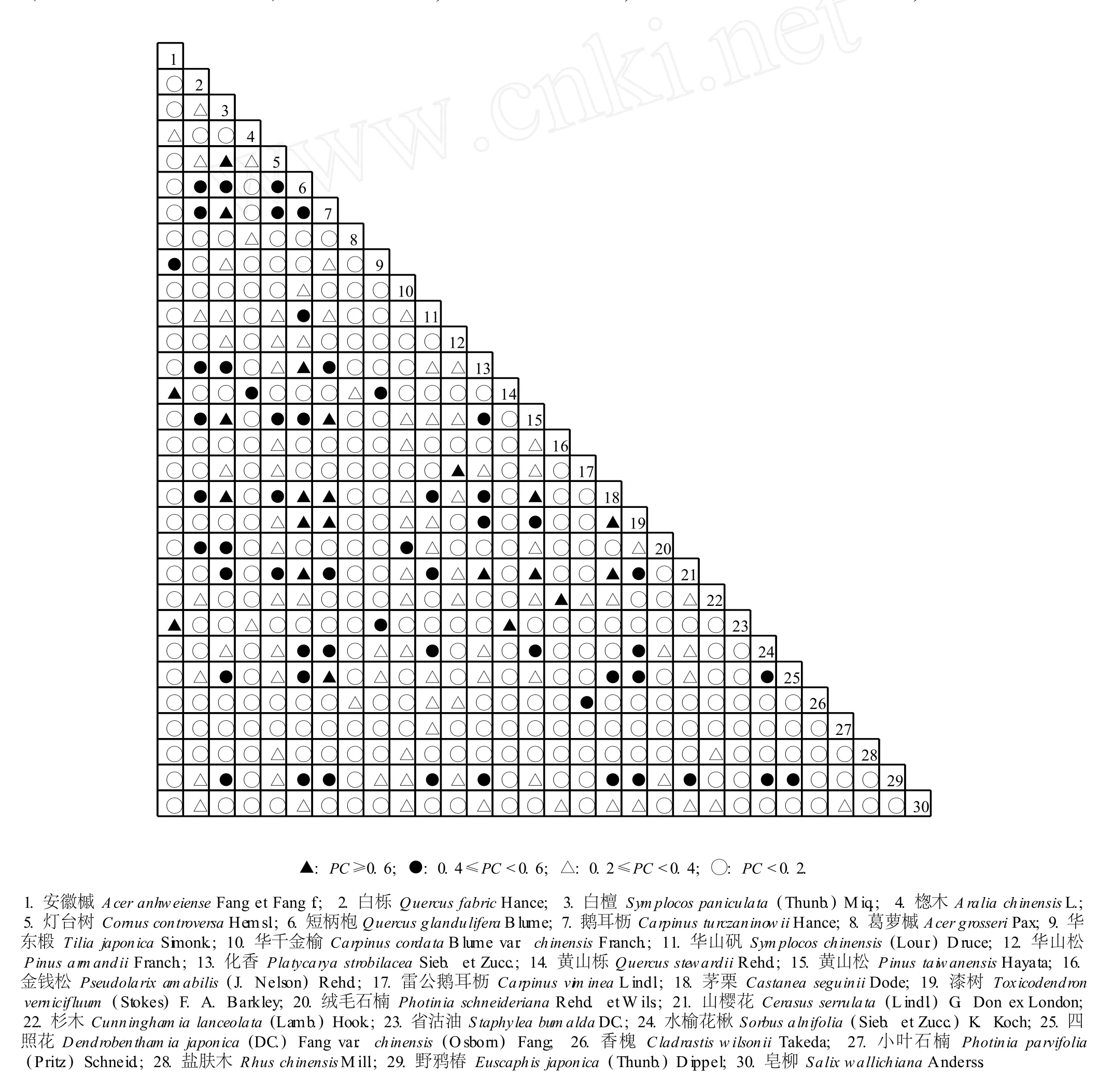
图 3 皖西大别山多枝尖山区主要乔木树种种间联结分析的共同出现百分率(PC)半矩阵图F ig.3 Percen tage of co-occurrence(PC)-sem im a tr ice d iagram of in ter-spec ific assoc ia tion ana lysis of dom inan tarbor spec ies in Duozh ijianm oun ta inous reg ion of Dab ieshan M oun ta in inW est Anhu i
皖西大别山多枝尖山区主要乔木树种中AC值在0.4以上 (AC≥0.4)的种对数仅占种对总数的 4.6%,表明该区域主要乔木种类中具有明显正联结性的种对较少;而具有明显负联结性的种对 (AC<-0.6)相对较多,占主要乔木种对总数的 26.0%;大多数乔木种类间的联结较为松散。
2.3 共同出现百分率(PC)分析
图 3结果表明:皖西大别山多枝尖山区主要乔木树种中共同出现百分率(PC)值在 0.6以上 (PC≥ 0.6)的种对有 22个,占主要乔木种对总数的 5.1%,代表种对有白檀与灯台树、鹅耳枥、黄山松和茅栗等;PC值介于 0.4~0.6(0.4≤PC<0.6)的种对有 53个,占主要乔木种对总数的 12.2%;PC值介于 0.2~0.4(0.2≤PC<0.4)的种对有 89个,占主要乔木种对总数的 20.5%;PC值低于 0.2(PC<0.2)的种对有271个,占主要乔木种对总数的 62.3%。与AC值反映的结果一样,PC值的分析结果也显示出皖西大别山多枝尖山区主要乔木种类中具有明显正联结性(PC≥0.6)的种对较少,大多数乔木种类间的联结较为松散。
3 讨 论
种间联结反映出物种间对生长环境综合生态因子反应的差异,2个物种的正联结程度越高说明它们所需的生长环境条件越相似;反之则表明对生长环境条件的需求有所不同[3]。然而,植物物种之间也存在直接或间接的相互影响[24-26],种间联结程度与生态位重叠值之间有较大的相关性,种间正联结程度越高,生态位重叠程度也越高[27]。在相同的生长环境和竞争条件下,生态习性相同或相似的物种往往共同出现在同一群落中,其种间联结表现为显著的正联结,这在前人的研究中已经得到了验证[5-16],本研究结果进一步证实了这一规律。例如,喜光树种短柄枹与山樱花以及华山松和雷公鹅耳枥,喜温暖湿润气候及深厚肥沃且排水良好土壤的鹅耳枥与漆树都表现出显著的正联结性;而鹅耳枥耐阴且喜中性土壤,香槐喜温暖而湿润的气候并耐干旱和寒冷,二者表现为显著的负联结性;喜中性偏阴及湿润气候的省沽油与喜光树种黄山松[28]的生态习性差别较大,二者表现为负联结性。另外,人为活动对植物群落内种间关系的形成也有一定的影响[28-30]。本研究中,金钱松和杉木均为人工混合种植,导致这 2个物种之间呈极显著的正联结性(P<0.01)。植物群落内物种间的种间关系与该群落所处的演替阶段密切相关,在群落的演替初期,物种间尚未形成一定的种间关系;演替中期,物种间主要表现为竞争关系;群落演替到达中生阶段时,物种间的关系较为复杂,群落内部处于同一层次的物种由于对生长环境需求相似而表现出种间联结[25-26]。本研究中,安徽槭、黄山栎和省沽油三者之间呈显著正联结,这3个物种都出现在海拔比较高的地段,受人为干扰的程度较低,植被较为原始;华山矾与水榆花楸、化香与山樱花以及短柄枹与山樱花则出现在原始植被遭到破坏后的次生林中,种间联结关系松散。由于本研究区域的植被曾遭到破坏,仅在部分人类难以到达的地段残留部分原始植被,植被的次生性可能是导致研究区域内植物种间联结松散的主要原因。
种间联结分析结果受到取样面积的影响。取样面积过大,可能导致正联结过多;取样面积过小,则又可能得出负联结过多的结果。因此,取样时应综合考虑研究区域内物种和生境的变化幅度,以确定最适宜的最小取样面积[8,12-13]。X2检验能够比较客观准确地判断出种对联结的显著性,共同出现百分率 (PC)和联结系数(AC)则能够判断出X2检验不显著的种对间的联结性及其大小。然而,虽然AC值和PC值能够反映出种间联结性的相对强弱,但其数值也受到取样面积及物种频度分布的影响,AC值和PC值的分类等级也没有统一的标准[12]。本研究中选取出现频率大于 5%的种类进行种间联结分析,在一定程度上可避免由于上述原因对种间联结分析结果造成的影响。综合上述原因,在对群落物种的种间联结研究中应以X2检验为基础,并结合AC值和PC值来比较植物群落内物种的种间联结特性,以获得更加准确的结果。
分析结果表明:皖西大别山多枝尖山区植物群落物种间的种间联结较为松散,由 30个主要乔木树种组成的 435个种对中有 15个种对的种间正联结性达到显著水平,其中极显著(P<0.01)正联结的种对有5个;负联结种对有 8个,其中极显著负联结的种对有2个。物种生态习性、群落演替阶段和人为活动等因素是导致本研究区域上述乔木物种联结特征的主要原因。研究结果揭示了该区域乔木优势种的相互关系,可为皖西大别山多枝尖山区植被恢复和管理提供借鉴和参考依据,同时也可为进一步研究其植被演替规律、维持和保护其物种多样性奠定基础。
致谢:在野外调查中得到了安庆师范学院生命科学学院李瑛、周光霞、贾名仙、刘振和晏伟伟等同学的帮助,特此致谢!
[1]PhillipsO L,Vargas PN,Monteagudo A L,et al.Habitat association among Amazonian tree species:a landscape-scale app roach [J].Journalof Ecology,2003,91:757-775.
[2]Gleason H A.The individualistic concep t of the p lant association [J].Bulletin of the Torrey BotanicalC lub,1926,53:7-26.
[3]王伯荪,彭少麟.南亚热带常绿阔叶林种间联结测定技术研究Ⅰ.种间联结测式的探讨与修正[J].植物生态学与地植物学丛刊,1985,9(4):274-285.
[4]张金屯.数量生态学[M].北京:科学出版社,2005.
[5]王乃江,张文辉,陆元昌,等.陕西子午岭森林植物群落种间联结性[J].生态学报,2010,30(1):67-78.
[6]邓贤兰,刘玉成,吴 杨.井冈山自然保护区栲属群落优势种群的种间联结关系研究[J].植物生态学报,2003,27(4): 531-536.
[7]郭志华,卓正大,陈 洁,等.庐山常绿阔叶、落叶阔叶混交林乔木种群种间联结性研究[J].植物生态学报,1997,21(5): 424-432.
[8]李德志,杨茂林.天然次生林群落中主要树木种群间联结关系的研究[J].植物生态学报,1996,20(3):263-271.
[9]骆土寿,李意德,陈德祥,等.海南岛鸡毛松人工林群落种间联结性研究[J].生态学杂志,2005,24(6):591-594.
[10]史作民,刘世荣,程瑞梅,等.宝天曼落叶阔叶林种间联结性研究[J].林业科学,2001,37(2):29-35.
[11]尹林克,李 涛.塔里木河中下游地区荒漠河岸林群落种间关系分析[J].植物生态学报,2005,29(2):226-234.
[12]张思玉,郑世群.福建永定桫椤群落内主要灌木种群的种间联结性研究[J].云南植物研究,2002,24(1):17-22.
[13]赵则海,祖元刚,杨逢建,等.东灵山辽东栎林木本植物种间联结取样技术的研究[J].植物生态学报,2003,27(3):396-403.
[14]胡 楠,范玉龙,丁圣彦,等.伏牛山自然保护区森林生态系统乔木植物功能型分类 [J].植物生态学报,2008,32(5): 1104-1115.
[15]杨晓东,吕光辉,张雪梅,等.艾比湖湿地自然保护区 8个乔灌木种群空间分布格局分析[J].植物资源与环境学报,2010, 19(4):37-42.
[16]邓福英,臧润国.海南岛热带山地雨林天然次生林的功能群划分[J].生态学报,2007,27(8):3240-3249.
[17]单人骅,刘昉勋.安徽省大别山区的植被及其地理分布纪要[J].植物生态学与地植物学丛刊,1964,2(1):93-102.
[18]刘 宾.安徽省大别山陀兴山区植物区系的研究[J].武汉植物学研究,1991,9(3):239-246.
[19]谢中稳,吴国芳.安徽大别山多枝尖山区植物区系的研究[J].华东师范大学学报:自然科学版,1993(1):102-110.
[20]高 林,徐如松,曹姣仙,等.鹞落坪国家级自然保护区植被类型的初步探讨[J].生物学杂志,2002,19(6):32-34,42.
[21]谢中稳,杜国华,张尽忠,等.安徽省鹞落坪自然保护区植物区系的数量分析[J].生物数学学报,1994,9(4):240-246.
[22]谢中稳,吴国芳.安徽省鹞落坪自然保护区植被类型及其分布[J].华东师范大学学报:自然科学版,1995(3):93-102.
[23]Yates F E,M arch D J,Iberall A S.Integration of the whole organism:a foundation for theoretical biology[M]∥Behnke JA. Changing B iological Problem s:D irections Toward Their Solution. New York:Oxford University Press,1972:110-132.
[24]彭李菁.鼎湖山气候顶极群落种间联结变化[J].生态学报, 2006,26(11):3732-3739.
[25]周先叶,王伯荪,李鸣光,等.广东黑石顶自然保护区森林次生演替过程中群落的种间联结性分析[J].植物生态学报, 2000,24(3):332-339.
[26]PetersH A.Neighbour-regulatedmortality:the influence ofpositive and negative density dependenceon tree populations in species-rich trop ical forests[J].Ecology Letters,2003,6:757-765.
[27]彭少麟,王伯荪.鼎湖山森林群落优势种群生态位重叠研究[M]∥中国科学院鼎湖山森林生态系统定位研究站.热带亚热带森林生态系统研究:第6集.北京:科学出版社,1990.
[28]《安徽植物志》协作组.安徽植物志:第 4卷[M].合肥:安徽科学技术出版社,1986.
[29]苏炳霖,何东进,洪 伟,等.武夷山风景名胜区天然林乔木层主要种群的种间联结性研究[J].植物资源与环境学报, 2007,16(3):40-45.
[30]苏小青,林思祖,黄石德.干扰状态下闽楠林乔木层主要种群种间联结性的研究[J].中国生态农业学报,2007,15(5):7-10.
