白眉山鹧鸪冬季觅食地选择
2011-12-25潘红平黄家喜
杨 岗, 潘红平, 许 亮, 周 放, 黄家喜
(广西大学 动物科学技术学院, 广西 南宁 530005)
白眉山鹧鸪冬季觅食地选择
杨 岗, 潘红平, 许 亮, 周 放*, 黄家喜
(广西大学 动物科学技术学院, 广西 南宁 530005)
2010年11—12月, 在广西九万山国家级自然保护区对白眉山鹧鸪冬季觅食地选择进行研究。发现觅食点34个, 设置利用样方和对照样方各25个, 测量并分析19个有关参数。结果显示, 白眉山鹧鸪对阔叶林、针阔混交林、阔叶摆竹混交林和摆竹林表现为随机利用, 对毛竹林和杉木林表现为避开选择;其偏向选择面朝东南方向的山坡, 觅食地坡度在20~44°之间, 觅食地摆竹灌丛密度和摆竹灌丛盖度、草本密度和草本盖度都显著低于对照样方, 落叶盖度显著高于对照样方。逐步判别分析结果表明, 落叶盖度、坡度和摆竹灌丛盖度是区分冬季觅食地和对照样方的主要参数, 判别准确率达86.0%。白眉山鹧鸪冬季对觅食地的选择与食物资源和躲避危险有关。因此, 建议在保护白眉山鹧鸪时应着重关注这两个因素。
白眉山鹧鸪; 觅食地选择; 判别分析
白眉山鹧鸪(Arborophila gingica), 又名白额山鹧鸪, 是中国南方山地和丘陵地区的特有鸟类(Lei & Lu, 2006)。IUCN(2010)将其列为易危物种。该鸟分布区域狭窄, 仅见于浙江西南部、福建西北部、江西南部、湖南南部、广东北部、广西东部和中部 (Wang & Xie, 2009; Zheng, 2005)。以往, 对白眉山鹧鸪的研究较少, 只有习性观察, 分布等部分报道(Li, 1990; Zhou, 1996; He et al, 2007)。
适宜的栖息地能为物种提供其生存、繁殖和种群延续的环境条件(Block & Brenan, 1993; Boyce &McDonald, 1999), 栖息地利用的评估是物种生态需求的一部分, 是进行物种有效保护的先决条件(Luck, 2002; Oppel et al, 2004)。觅食地是栖息地的重要组成, 对觅食地选择的研究能够在众多环境因子中找出影响动物选择的关键因子, 并了解其选择机制, 从而有助于动物的保护和种群恢复。2010年11—12月我们对广西九万山国家级自然保护区的白眉山鹧鸪冬季觅食地选择进行定量研究。
1 研究地区与方法
1.1 研究地区
广西九万山国家级自然保护区(E108°35′32″~108°48′49″, N25°01′55″~25°19′54″)位于广西壮族自治区的北部。地貌类型属于侵蚀褶皱深~中等切割中山, 相对高差900~1 200 m, 最高海拔1 693 m; 年平均气温12.0~17.1℃, 最冷1月平均气温4.0~6.0℃, 极端最低气温−8.0℃, 年降水量1 600~2 100 mm,年均相对湿度82%~90%, 无霜期303 d, 12月中旬—次年2月中旬为霜冻期; 保护区地表水充足, 溪河纵横交错。
本研究在保护区的清水塘站和杨梅坳站进行, 海拔1 300 m以下植被为中亚热带典型常绿阔叶林, 主要树种有米槠(Castanopsis carlesii)、红锥(Castanopsis hystrix)、贵州锥(castanopsis kweichowensis)、红背甜槠(Castanopsis neocavaleriei)、银木荷(Schima argentea); 海拔1 300~1 500 m为中山常绿落叶阔叶混交林和中山针叶阔叶混交林, 主要树种有钩锥(Castanopsis tibetana)、水青冈(Fagus longipetiolata)、小叶罗汉松(Podocarpus wangii)、光枝杜鹃(Rhododendron haoful); 大于1 500 m的山顶和山脊地段出现有矮林, 主要有毛棉杜鹃(Rhodo dendron moulmainense)、多花杜鹃(Rhododendron cavaleriei)、猴头杜鹃(Rhododendron simiarum); 常绿阔叶林、常绿落叶阔叶混交林和山顶矮林中镶嵌有小块或小片摆竹(Indosasa shibatae- oides); 人工林主要分布在400~900 m, 有杉木林(Cunninghamia lanceolata)和毛竹林(Phyllostachys edulis)(Ning et al, 2010)。
1.2 研究方法
1.2.1 数据收集 2010年 11—12月, 在当地霜冻期前对白眉山鹧鸪冬季觅食地选择进行研究。研究区内同域分布有白眉山鹧鸪和白鹇(Lophura nythemera)两种雉类, 白眉山鹧鸪拨开落叶层取食, 呈小片取食痕迹; 白鹇的取食痕迹为锥形的取食坑, 深度 5~8 cm。搜寻白眉山鹧鸪的取食痕迹, 用 GPS记录取食点所在位置, 如果邻近有多个取食点(相距<5 m), 则以多个取食点的中心为取食位点, 当两个取食位点之间的直线距离大于100 m时, 以取食位点为中心, 设定10 m×10 m的大样方作为利用样方, 在大样方中心和四角设置5个1 m×1 m小样方(Young et al, 1991)。以大样方测度地形、乔木层、摆竹灌丛层等信息, 以小样方测度草本层和落叶层信息, 测量觅食地参数19个:植被类型、海拔、坡向、坡位、坡度、与林缘距离、与林间小路距离、与水源距离、乔木密度、乔木均高、乔木盖度、摆竹灌丛密度、摆竹灌丛均高、摆竹灌丛盖度、草本密度、草本均高、草本盖度、落叶盖度以及落叶厚度。植被类型分为阔叶林、针阔混交林、阔叶摆竹混交林、摆竹林、毛竹林以及杉木林。坡向是指取食点所在山坡正对的方向, 取值通过指北针测定,为自正北方向顺时针旋转至山坡正对方向时指针所转过的度数。坡位是指取食点所在山坡的位置,分为下坡位、中坡位和上坡位, 取值分别为1、2、3(Xu et al, 2002)。以利用样方的中心为参照点, 通过随机数字表确定对照样方的中心(Brennan et al,1987), 设置相同面积的对照样方, 在对照样方中测量与利用样方相同的参数。
1.2.2 数据处理 使用 Bailey’s方法(Bailey, 1980)分析白眉山鹧鸪对不同植被类型的选择性, 不需要预先进行拟合卡方检验(Cherry, 1998)。如果某种植被类型的期望利用比例在 Bailey’s置信区间之内,表示对该植被类型随机利用; 如果在置信区间的左侧或右侧, 表示正选择或负选择。植被类型的期望利用比例为研究区内某种植被类型的面积占总面积的百分比, 对某种植被类型的实际利用比例为在这种植被类型中觅食点的数量占所有觅食点数量的百分比。用圆形分布统计方法(circular distribution)分析坡向和坡度(Zar, 2010); 采用
Rayleigh’s test 分析坡向和坡度是否有集中趋势,若存在集中趋势, 则采用求平均角期望区间的方法得出白眉山鹧鸪对坡向和坡度的选择性; 采用
Watson’sU2test 分析坡向和坡度在利用样方和对照样方之间是否存在差异。分析其余变量在利用样方和对照样方之间的差异时, 先用 Kolmogorov-Smirnov test 检验数据是否符合正态分布。当数据符合正态分布时, 使用独立样本的t-test; 当变量不符合正态分布时, 使用Mann-Whitney test。利用样方和对照样方之间的变量存在显著差异时, 用Spearman correlation 判别变量之间的相关性, 当两变量之间的相关系数r的绝对值≥0.60时, 则取生物学意义比较重要的变量进入下一步分析 (Lahaye& Gutierrez, 1999)。采用逐步判别分析(stepwise discriminant analysis)对剩余觅食地变量进行分析,以确定影响白眉山鹧鸪觅食地选择的关键因子。使用逐步判别分析时, 所有选项均为系统默认值。数据采用Mean±SD表示, 其中Mean为算术平均值,SD为标准差。
数据处理均在 Microsoft Excel 2003和 SPSS 17.0 for Windows统计软件上完成。
2 结 果
2.1 植被类型
记录觅食点34个, 阔叶林内18个, 针阔混交林内7个, 阔叶摆竹混交林内5个, 摆竹林内4个。测量利用样方和对照样方各 25个。白眉山鹧鸪随机利用阔叶林、针阔混交林、阔叶摆竹混交林和摆竹林, 而回避毛竹林和杉木林(表1)。
2.2 地形因素
对利用样方的分析表明, 觅食生境的坡向存在集中趋势(Z=6.991,Z0.01,25=4.483,P<0.01), 其平均角为124° (95%置信区间为94~154°),白眉山鹧鸪偏向选择东南坡; 觅食点的坡度也存在集中趋势(Z=24.301,Z0.01,25=4.483,P<0.01), 其平均角为 32°(95%置信区间为 20~44°), 白眉山鹧鸪选择有一定坡度的生境。

表1 白眉山鹧鸪对不同植被类型的选择性Tab. 1 Habitat availability and actual use by White-browed Hill Partridge in Jiuwanshan, Guangxi
与对照样方的坡度和坡向的检验结果表明, 白眉山鹧鸪对坡向的选择并不显著(U2=0.183,U20.05,25,25=0.185,P>0.05), 而对坡度具有极显著的选择性(U2=0.296,U20.01,25,25=0.262,P<0.01)。
2.3 植被因素
白眉山鹧鸪的利用样方和对照样方相比, 在摆竹灌丛密度、摆竹灌丛盖度、草本密度、落叶盖度等4个参数上存在极显著差异, 在草本盖度上存在显著差异(表2)。在利用样方中, 摆竹灌丛密度和草本密度较低, 盖度较小, 落叶盖度较大。
2.4 影响白眉山鹧鸪觅食地选择的主要因子
将表2中差异显著和极显著的5个变量以及坡度进行 Spearman相关系数分析, 检验变量之间是否相互独立, 发现摆竹灌丛盖度和摆竹灌丛密度,以及草本密度和草本盖度之间的相关系数绝对值>0.60(r分别为0.821和0.630)。因此, 剔除摆竹灌丛密度和草本密度, 将其余4个变量纳入逐步判别分析, 筛选白眉山鹧鸪觅食地选择的关键因子(表3)。从表中可以看出, 在区分利用样方和对照样方时, 根据贡献值的大小, 依次为落叶盖度、坡度和摆竹灌丛盖度。标准化的典型判别函数为Z=−0.651×落叶盖度+0.625×坡度+0.539×摆竹灌丛盖度(Z<0,为利用样方; Z>0,为对照样方), 这3个变量构成的方程在对利用样方和对照样方进行区分时, 正判率可以达到86.0%。
3 讨 论
据我们观察, 冬季白眉山鹧鸪主要扒开落叶层觅食, 取食落叶层下的植物种子和小动物。与Li et al (1990)观察冬季白眉山鹧鸪食物主要包括有红栲、野漆、乌药等植物种子和果实, 蚂蚁、虫蛹和小螺丝等动物性食物的结果相同。实地调查发现,有一定落叶盖度的区域, 土壤湿度更高, 含水量更大, 土壤动物更丰富, 一些果实和种子也在落叶层中。白眉山鹧鸪避开选择植被单一, 提供食物有限的杉木林和毛竹林, 而倾向于选择阔叶林, 这与阔叶林能提供更多的果实和其他食物有关。白眉山鹧鸪偏向选择向阳的东南坡, 因为当地冬季气温相对较低, 向阳坡能提供较温暖的微环境气候, 一方面减轻白眉山鹧鸪抵御寒冷天气的能量消耗; 另一方面温暖的环境也更适宜土壤动物活动。其在坡度上倾向选择20~44°的中缓坡, 坡度陡峭的坡面落叶盖度较低, 而有落叶覆盖的区域枯叶松散且较厚,对其扒食不利。判别分析结果表明, 与食物资源有关的落叶盖度是其觅食地选择的关键因子, 且权重最大(表3)。因此, 可认为白眉山鹧鸪在觅食地选择上首先考虑食物因素。

表2 冬季白眉山鹧鸪不同尺度利用样方和对照样方之间的变量比较 (Mean±SE)Tab. 2 Comparisons of variables between the White-browed Hill Partridge used sites and control sites during winter in Jiuwanshan, Guangxi (Values are Mean±SE)
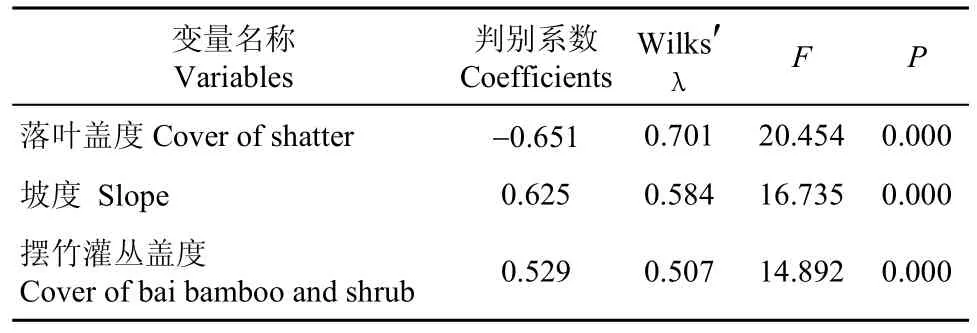
表3 白眉山鹧鸪利用样方和对照样方变量的逐步判别分析结果Tab. 3 Consequences of Stepwise Discriminant Analysis of the White-browed Hill Partridge habitat variable between used sites and control sites during winter in Jiuwanshan, Guangxi
白眉山鹧鸪避开选择摆竹灌丛层和草本层较密的生境是由于中下层植被茂盛的生境将影响其觅食活动和转移, Li et al (2003)也指出四川山鹧鸪(Arborophila rufipectus)选择林下空隙较大的生境。对一些雉类的研究表明, 与隐蔽性有关的因子是雉类栖息地选择的关键因子(Li et al, 2010; Liu et al,2008), 白眉山鹧鸪选择草本盖度、摆竹灌丛层盖度较低的生境, 虽然在一定程度上降低了其隐蔽性,但有利于其观察周围环境中的潜在威胁; 而且白眉山鹧鸪与其他大型雉类相比, 体型较小, 活动能力也较强, 当发现危险时, 能快速逃离。冬季时常有农户进山採集菌类, 较低的摆竹灌丛盖度能让白眉山鹧鸪在较远处时即可发现人类, 并进行躲避反应。调查时曾两次惊飞白眉山鹧鸪, 距离数十米,其应该是在林下穿行一段距离后被迫飞离。白眉山鹧鸪的连续扒食点较少, 可能是由于食物资源少,而减少觅食投入;也可能是为避免在同一地点长时间逗留, 从而降低被天敌发现的机率。白眉山鹧鸪觅食地选择的策略应是权衡觅食收益与防范被捕食及其他干扰因素后的综合考量。
调查区域内, 部分地区摆竹生长旺盛, 已成为灌木层的优势植被, 影响其他灌木生长, 也影响阔叶林幼苗的发育, 其单一的优势种群将会降低植被的多样性, 是多样性稳定的不利因素。摆竹林镶嵌在阔叶林内, 植被相对单一, 食物资源并不丰富,白眉山鹧鸪选择在其中觅食, 可能是对摆竹扩散长期适应的耐受结果, 也可能是由于阔叶林内竞争压力过大而被迫选择, 摆竹林的扩张对白眉山鹧鸪的影响值得进一步研究。从觅食点记录可看出阔叶林,针阔混交林, 阔叶摆竹混交林、摆竹林的觅食点次数是逐渐递减的, 为了维持白眉山鹧鸪种群的长期稳定和发展, 应注意控制保护区内摆竹的生长。冬季农户进山採集野生食用菌活动频繁, 活动范围较广, 干扰白眉山鹧鸪的正常觅食, 而冬季是白眉山鹧鸪为即将到来的繁殖季做准备的时期, 良好的生存环境有利于种群存活和种群繁衍, 应减少人为活动的干扰。
致谢:广西九万山国家级自然保护区领导给予本研究大力支持, 野外工作得到清水塘站和杨梅坳站全体工作人员和当地群众的帮助, 在此一并表示衷心感谢!
Bailey BJR. 1980. Large sample simultaneous confidence intervals for the multinomial probabilities based on transformation of the cell frequencies [J].Technometrics, 22: 583-589.
Block, WM, Brenan, LA. 1993. The habitat concept in ornithology, theory and applications [J].Curr Ornith, 11: 35–91.
Boyce, MS, McDonald L. 1999. Relating populations to habitats using resource selection functions [J].Tree, 4 (7): 268–272.
Brennan LA, Block WM, Gutibrrez RJ. 1987. Habitat use by mountain quail in northern California [J].Condor, 89, 66–74.
Cherry S. 1998. Statistical tests in publications of The Wildlife Society[J].Wildl Soc Bull, 26(4): 947-953.
He FQ, Lin Z, Jiang HD, Lin JS. 2007. The occurrence of the White-necklaced Partridge in Fujian of SE China [J].Chn J Zool, 42(4):147-148. [何芬奇, 林植, 江航东, 林剑声. 2007. 白眉山鹧鸪在福建的分布. 动物学杂志, 42(4): 147-148.]
IUCN. 2010. The IUCN Red List of Threatened Species [EB/OL].http://www.iucnredlist.org/apps/redlist/search.
Lahaye WS, Gutierrez RJ. 1999. Nest sites and nesting habitat of the Northern Spotted Owl in northwestern California [J].Condor, 101:324-330.
Lei FM, Lu TC. 2006. China endemic Birds [M]. Beijing: Science Press,76-79. [雷富民, 卢汰春. 2006. 中国鸟类特有种. 北京: 科学出版社,76-79.]
目前的图形密码,组成图形均为3*3的正方形(下文称“3*3式”),在密码的规则上,也均为在3*3图形中九个点之间相互连线。此种图形密码所包含的元素点仅9个,规则也较单一,转换成的密文过于简易,被密钥解析法破解时所用时间也较短。另外,图形密码目前为止只有3*3式应用较广,其余的图形几乎没有被尝试。除数字密码和3*3式图形密码安全性的比较以外,将3*3式与其他图案在安全性上相比较也是必要的。基于上述考虑,研究以下问题:
Li C, Hu J, Yu ZW. 2003. Distribution and Habitat Selection of the Sichuan
Hill-Partridge [J].Chn J Zool, 38(6): 46-51. [李操, 胡杰, 余志伟.2003. 四川山鹧鸪的分布及生境选择. 动物学杂志, 38(6): 46-51.]
Li XH, Tan HZ, Chen CA, Zhang AL. 1990. Ecological observation on White-browed Hill Partridge [J].Chn J Zool, 25(1): 22-24. [李晓惠,谭洪治, 陈采安, 张爱理. 1990. 白额山鹧鸪的生态观察. 动物学杂志, 25(1): 22-24.]
Li W, Zhou W, Liu Z, Li N. 2010. Habitat selection change of Mrs. Hume Pheasant (Syrmaticus humiae) in Dazhongshan during the year [J].Zool Res, 31(5): 499-508. [李伟. 周伟, 刘钊, 李宁. 云南大中山黑颈长尾雉栖息地选择周年变化. 动物学研究, 31(5): 499-508.]
Luck, GW. 2002. The habitat requirement of the Rufous Treecreeper(Climacteris rufa). 1. Preferential habitat use demonstrated at multiple spatial scales [J].Biol Conserv. 105: 383–394.
Liu Z, Zhou W, Zhang RG, Xie YC, Huang QW, Wen YY. 2008. Foraging sites selection of green peafowl (Pavo muticus imperator) in different seasons in Shiyangjiang Valley of upper Yuanjiang drainage, Yunnan[J].Biodiv Sci, 16(6): 539-546.[刘钊, 周伟, 张仁功, 谢以昌, 黄庆文, 文云燕. 2008. 云南元江上游石羊江河谷绿孔雀不同季节觅食地选择. 生物多样性, 16(6): 539-546.]
Ning SJ, Su Y, Tan XF. 2010. Integrated Investigation Report on Jiuwanshan National Nature Reserve, Guangxi [M]. Beijing: Science Press , 86-227.[宁世江, 苏勇, 谭学锋. 2010. 生物多样性关键地区-广西九万山自然保护区科学考察集. 北京: 科学出版社, 86-227.]
Wang S, Xie Y. 2009. China Species Red List vol.Ⅱ Vertebrates Part 2 [M].Beijing: Higher Education Press, pp185. [汪松, 解焱. 2009. 中国物种红色名录(第 2卷): 脊椎动物(下册). 北京: 高等教育出版社,185.]
Xu JL. Zhang XH, Zhang ZW, Zheng GM. 2002. Brood habitat characteristics of Reeve’S Pheasant (Syrmaticus reevesii) in Dongzhai National Nature Reserve[J].Zool Res, 23(6): 471-476. [徐基良, 张晓辉, 张正旺, 郑光美. 2002. 白冠长尾雉育雏期的栖息地选择. 动物学研究, 23(6): 471-476.]
Young L, Zheng GM, Zhang ZW. 1991. Winter movements and habitat use by Cabot’s Tagopans in Southeastern China [J].Ibis, 133: 121-126.
Zar JH. 2010 . Biostatistical Analysis[M]. Fifth Edition . New Jersey, USA:Prentice Hall, Inc. 605-668.
Zheng GM. 2005. A Checklist on the Classification and Distribution of the Birds of China [M]. Beijing: Science Press, 53. [郑光美. 2005. 中国鸟类分类与分布名录. 北京: 科学出版社, 53.]
Zhou F. 1996. The distribution of White-browed Hill Partridge(Arborophila gingica) in Guangxi [J].Acta Zool Sin, 42: 143. [周放.1996. 广西白额山鹧鸪的分布. 动物学报, 42: 143.]
Winter feeding sites selection of White-browed Hill Partridge
YANG Gang , PAN Hong-Ping , XU Liang , ZHOU Fang*, HUANG Jia-Xi
(College of Animal Science and Technology,Guangxi University,Nanning530005,China)
The winter feeding sites of White-browed Hill Partridge (Arborophila gingica) was investigated in Jiuwanshan National Nature Reserve in Guangxi from November to December 2010. With the 34 found feeding sites, the used sites (n=25) were compared with the control sites (n=25), and 19 parameters were measured at each site. The results showed that White-browed Hill Partridge randomly use broadleaved forest, mixed coniferous-broadleaved forest, mixed broadleaved-bai bamboo (Indosasa shibataeoides) forest and bai bamboo forest, while they rarely occurred in mao bamboo (Phyllostachys edulis) forest and China fir (Cunninghamia lanceolata) forest. The birds prefer to select the southeast-facing slops of 20 − 44 degrees, with smaller bai bamboo-shrub and grass density, lower bai bamboo-shrub and grass coverage, and greater shatter coverage. We found that the main selection factors at the used and control feeding sites, using the Step DA, were the shatter coverage, slope, and bai bamboo-shrub coverage. The veracity to distinguish was 86.0%. The feeding sites selection of White-browed Hill Partridge is associate with food resources and safety index,thus, we suggest that the protection should focus on these two aspects.
White-browed Hill Partridge (Arborophila gingica); Feeding-site selection; Step discriminant analysis
Q959.725; Q958.11
A
0254-5853-(2011)05-0556-05
10.3724/SP.J.1141.2011.05556
2011-03-03; 接受日期:2011-07-22
∗通讯作者(Corresponding author), E-mail: zhoufang@gxu.edu.cn
男, 硕士研究生, 研究方向为动物资源保护。E-mail: wild_bird@126.com
