SCG10基因重组质粒的构建及蛋白表达和定位
2011-12-18刘芙蓉李彦姝张红艳苏楠李丰
刘芙蓉 李彦姝 张红艳 苏楠 李丰
SCG10基因重组质粒的构建及蛋白表达和定位
刘芙蓉 李彦姝 张红艳 苏楠 李丰*
(中国医科大学卫生部细胞生物学重点实验室,医学细胞生物学教育部重点实验室,细胞生物学教研室,辽宁沈阳110001)
目的 构建SCG10真核表达载体并证实融合蛋白在细胞内表达及定位。方法 以人胎脑cDNA文库为模板,PCR扩增SCG10全长编码基因,亚克隆至p EGFP-C1表达载体中。将构建的重组质粒测序并转染到人胚肾 HEK293中,提取细胞蛋白进行Western blot检测。利用共聚焦激光扫描显微镜观察p EGFP-SCG10在 HEK293细胞内定位。结果SCG10全长基因序列克隆到了真核表达载体p EGFP-C1中,酶切鉴定片段大小540bp。Western blot检测到了融合蛋白表达,分子量约为48kD。p EGFP-SCG10在细胞内定位以细胞浆为主,在细胞核少量表达。结论 成功构建了SCG10全长基因真核表达载体,p EGFP-SCG10蛋白主要定位于 HEK293细胞浆内。
SCG10; Western blot; 质粒构建
SCG10(Superior cervical ganglion-10)是磷酸化蛋白stathmin家族成员之一,在脑组织发育[1]、微管运动和信号转导中均有重要的作用,在神经元生长和骨形成中也有调节作用。SCG10也称stathmin2,是特异的神经元标志蛋白和潜在的微管失稳因子[2]。虽然在成人脑组织中SCG10的表达水平低,但在发育的中枢神经系统中SCG10高度表达。
SCG10编码序列全长561bp,187个氨基酸,N-末端的定向序列调节它向轴突和树突生长点的运输,主要蓄积于生长锥中心区域(该区域含大量动态微管)。SCG10是通过神经元生长锥的微管失衡活化来调节神经元生长的主要调节子,能增加轴突和树突顶端的能动结构。轴突的延伸、分支和生长锥的伸展是随细胞骨架蛋白连续重组来完成的,其中微管是重要的组成部分。生长锥的微管末端随α-和β-微管蛋白异二聚体的聚合或解聚而持续的延伸或缩短[3]。SCG10通过分离αβ-微管蛋白异二聚体增加微管的动态不稳定性,从而降低聚合程度、增加突变频率反应和解聚微管[4]。
本实验旨在获得SCG10全长基因序列及其在细胞内的表达和定位,为进一步探讨它的生物学效应打下基础。
材料和方法
1.菌株、细胞、质粒及主要试剂
大肠杆菌DH5感受态为本实验室制备;HEK 293人胚肾细胞系为本实验室保存;质粒p EGFPC1(Clontech);PryobestTMDNA polymerase、dNTP、DNA电泳凝胶回收试剂盒、反转录试剂、限制性核酸内切酶 Eco R I和Bam H I(大连 Ta Ka-Ra);TRIzol RNA提取试剂(Invitrogen);DNA marker、Protein marker(GenScript);T4DNA 连接酶(NEB);anti-GFP抗体(GenScript);HRP标记的羊抗鼠 Ig G(Promega);引物合成、DNA序列测定由上海英俊生物有限公司完成;ECL发光试剂盒(Pharmacia);其他试剂为国产分析纯。
2.SCG10全长基因扩增
设计SCG10扩增的 PCR引物,并在引物中加入了 Eco R I和Bam H I两个限制性酶切位点。上游 SCG10引物为 5’-GCACA TGAA TTCAA TGGCTAAAACA GCA-3’,下游 SCG10 引物为 5’-GCA GCA GGA TCCTCA TTGCTTCTCTCC-3’。以人胎脑cDNA文库为模板PCR扩增SCG10全长编码序列。
3.p EGFP-SCG10表达载体的构建和瞬时转染
将p EGFP-C1载体和SCG10 PCR片段分别用Eco R I和Bam H I双酶切后,凝胶回收纯化产物。将两个片段用 T4DNA连接酶常温连接2h,取5μl连接产物转化到DH5α大肠杆菌感受态中,37℃培养过夜。挑取单克隆菌落,接种于含卡那霉素的LB培养基中,37℃振荡过夜。用碱裂解法提取质粒DNA,用 Eco R I和Bam H I双酶切鉴定外源基因的插入,进行测序分析。
HEK 293细胞用DMEM高糖培养基培养,细胞铺于六孔板至70-80%融合,按Invitrogen公司提供的LipofectamineTM2000转染六孔板的说明书进行操作。
4.蛋白质提取与Western blot鉴定
转染48h后,弃掉培养基,用 PBS清洗两遍。在细胞中加入60μl含有蛋白酶抑制剂的 RIPA裂解液,用橡皮刮刀将细胞缓慢刮下。收集所有液体到新的离心管中,冰上放置 10min,裂解细胞。13 000×g 4℃离心30min,将上清转到新的离心管中,取上清5μl进行定量。
将蛋白定量后,取 40μg总蛋白经 10%SDSPA GE凝胶分离,4℃过夜恒压转移至 PVDF膜上,5%脱脂奶粉封闭3h,TBST洗膜15min,3次。用anti-GFP抗体(1∶500稀释),4℃过夜,TBST洗膜15min,3次。再用辣根过氧化物酶标记的羊抗鼠(1∶5000稀释)二抗孵育 1h,TBST洗膜 15min,3次。ECL显色,压片。
5.共聚焦激光扫描显微镜观察p EGFP-SCG10细胞内定位
将构建的绿色荧光蛋白表达载体p EGFPSCG10转染到 HEK 293细胞24h后,应用OL YMPUS FV 1000S-SIM/IX81激光共聚焦扫描显微镜进行扫描,激发光波长为 488nm,可看到 GFPSCG10蛋白在细胞内的定位处呈现绿色。
结 果
1.人SCG10基因全长PCR扩增
以人胎脑cDNA文库为模板,特异性引物进行聚合酶链反应扩增 SCG10全长。扩增的条件为95℃5min预变性;95℃1min,55℃1min,72℃1min此三个温度进行30个循环;最后72℃10min,温度降至4℃。1%水平琼脂糖凝胶电泳,溴化乙啶染色后,在紫外灯下观察结果(图1)。

图1 PCR扩增出的 SCG10的全长序列.1.PCR DNA Marker;2,3.SCG10 PCR扩增产物Fig.1 PCRamplificationof thefull length SCG10.Lane1:PCRDNA Marker;Lane 2,Lane 3:PCRproductsof SCG10
2.SCG10重组真核表达质粒的构建和鉴定
将重组质粒 p EGFP-SCG10经 Eco R I和Bam H I双酶切后得到了约4.7 kb和540bp左右的两条带(图2)。经过测序分析,外源序列在NCBIBlast比对结果与SCG10编码序列一致。

图2 对融合质粒p EGFP-SCG10进行限制性酶切分析.1.PCR DNA Marker;2.Eco R I和Bam H I对融合质粒p EGFP-SCG10进行酶切鉴定;3,4.p EGFP-C1质粒;5.Lambda DNA/Eco RI+HindⅢMarkerFig.2 Restriction enzyme digestion analysis of the recombinant plasmid p EGFP-SCG10.Lane 1:PCR DNA Marker;Lane 2:Recombinant plasmid p EGFP-SCG10 digested by Eco R I和Bam H I;Lane 3,Lane4:p EGFP-C1 vectors;Lane 5:Lambda DNA/Eco RI+HindⅢMarker
3.真核表达质粒在 HEK 293细胞中的表达
将构建的真核表达质粒p EGFP-SCG10转染到HEK 293人胚肾细胞系中,48h以后将提取的蛋白经10%SDS-PA GE凝胶电泳,Western blot检测到了融合绿色荧光蛋白 GFP的SCG10蛋白表达(图3)。分子量约48kD,为质粒本身的 GFP标签分子量(27kD)和SCG10分子量(21kD)之和,说明融合蛋白表达。
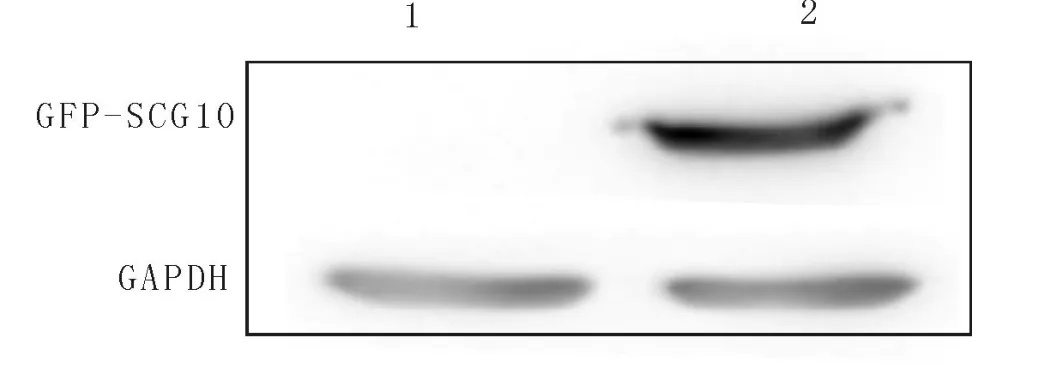
图3 用α-GFP鼠的单克隆抗体对融合蛋白 GFPSCG10的表达进行 Western blot检测.1.转染 HEK293细胞的 GFP蛋白;2.转染 GFP-SCG10的融合蛋白Fig.3 Western blot analysis of GFP-SCG10 f usion protein using anti-α-GFP mAB.Lane 1:HEK293 cells with GFP-control protein;Lane 2:The fusion protein of GFPSCG10
4.GFP-SCG10在细胞内的定位
通过共聚焦激光扫描显微镜观察到 GFP-SCG10在细胞内定位以浆为主,细胞核内少量表达(图4)。

图4 GFP-SCG10在 HEK293细胞内定位.A:GFP-SCG10(绿色);B:Topro 3(蓝色);C:图像叠加Fig.4 The localization of GFP-SCG10 in HEK293 cells.A:GFP-SCG10(green);B:Topro 3(blue);C:mergeof images
讨 论
磷酸化蛋白stathmin家族有4个蛋白分子,主要功能是通过抑制微管聚合、促进微管解聚来调节微管动力学。SCG10作为家族的一员,自身结构上有三个功能区域,分别是N末端的膜结合区域、中心调节区域和c末端的卷曲螺旋区域。N末端含有35个氨基酸,是家族中其他成员所不具有的,对其在高尔基体定位及在生长锥的富集起着至关重要的作用[5]。SCG10有四个丝氨酸磷酸化位点[6],在体内外,各种蛋白激酶能使这些位点发生磷酸化。中心调节区域有c AMP依赖蛋白激酶(PKA)催化S50和S97发生磷酸化,调节微管蛋白的解聚。c末端的卷曲螺旋区域被认为是蛋白的相互作用区域,也是SCG10与微管蛋白和细胞骨架相互结合的区域。SCG10蛋白在中枢神经系统和周围神经系统中同时表达。基因表达的下降与唐氏综合征[7]和阿尔茨海默氏病[8]的发生相关。
本实验以SCG10基因的上下游序列设计两对特异性的引物,并在引物中掺入了限制性酶切位点。利用引物从人胎脑cDNA文库中扩增出了SCG10的全长基因序列,并将其构建到了真核细胞表达载体p EGFP-C1中,测序分析外源基因序列,比对结果证实质粒构建成功。将融合质粒转染到 HEK 293细胞中,Western blot验证融合蛋白在细胞中表达。利用共聚焦激光扫描显微镜证实,p EGFPSCG10在 HEK 293细胞内定位以细胞浆为主,在细胞核内少量表达。基因全长的获得及其在细胞内的表达与定位,为进一步研究SCG10的生物学特性奠定了初步基础。
[1]Bieche I,Maucuer A,Laurendeau I,et al.Expression of stathmin family genes in human tissues:non-neuralrestricted expression for SCL IP.Genomics,2003,81:400-410
[2]Li Y.H,Ghavampur S,Bondallaz P,et al.Rnd1 regulates axon extension by enhancing the microtubule destabilizing activity of SCG10.J Biol Chem,2009,284:363-371
[3]Mitchison T,Kirschner M,Dynamic instability of microtubule growth.Nature,1984,312:237-242
[4]Sobczak A,Debowska K,Blazejczyk M,et al.Calmyrin1 binds to SCG10 protein(stathmin2)to modulate neurite outgrowth.Biochim Biophys Acta,2011,1813:1025-1037
[5]Togano T,Kurachi M,Watanabe M,et al.Role of Ser50 phosp horylation in SCG10 regulation of microtubule depolymerization.J Neurosci Res,2005,80:475-480
[6]Antonsson B,Kassel D.B,Di Paolo G,et al.Identification of in vitro phosphorylation sites in the growth cone protein SCG10.Effect Of phosphorylation site mutants on microtubule-destabilizing activity.J Biol Chem,1998,273:8439-8446
[7]Bahn S,Mimmack M,Ryan M,et al.Neuronal target genes of the neuron-restrictive silencer factor in neurospheres derived from fetuses with Down’s syndrome:a gene expression study.Lancet,2002,359:310-315
[8]Okazaki T,Wang H,Masliah E,et al.SCG10,a neuron-specific growth-associated protein in Alzheimer’s disease.Neurobiol Aging,1995,16:883-894
Construction of recombinant plasmid of SCG10 gene and identif ication of
its fusion protein expression and localization
Liu Furong,Li Yanshu,Zhang Hongyan,Su Nan,Li Feng*
(Department of Cytobiology,Key L aboratory of Cytobiology,Ministry of Public Health,and Key L aboratory of Medical Cytobiology,Ministry of Education,China Medical University,Shenyang 110001,China)
Objective To construct the recombinant plasmid of SCG10 gene and identif y its f usion protein expression and localization.Methods The SCG10 coding sequence was amplified by polymerase chain reaction(PCR),and was subcloned into the p EGFP-C1 vector.After the target region was sequenced,the plasmid was transfected into HEK 293 cell lines.The expression of the recombinant plasmid in HEK 293 cells was proved by Western blot.The localization of p EGFP-SCG10 in HEK 293 was observed by using laser scanning confocal microscopy.Results SCG10 was successf ully constructed into the expressing vector p EGFP-C1.The length of the f ragment was 540bp,identified by restriction enzymes digestion.The expression of GFP-SCG10 f usion protein was detected by Western blot,with a molecular weight of 48kD.The p EGFP-SCG10 protein was mainly localized in the cytoplasm,less in the nucleus.Conclusion The recombinant plasmid was successf ully cloned into the eukaryotic expressing vector.The p EGFP-SCG10 f usion protein was expressed mainly in the cytoplasm.
SCG10; Western blot; Plasmid construction
R329.2 Q257
A
10.3870/zgzzhx.2011.03.014
2011-02-14
2011-04-27
国家自然科学基金(30771128;90813038);辽宁省教育厅创新团队项目(2008T195)
刘芙蓉,女 (1967年),汉族,学士。
*通讯作者(To whom correspondence should be addressed)
猜你喜欢
——水芹主要害虫识别与为害症状
