密度对泽陆蛙蝌蚪生长发育和变态的影响
2011-11-23施林强马小梅
施林强,毛 飞,马小梅
(杭州师范大学生命与环境科学学院,浙江 杭州 310036)
密度对泽陆蛙蝌蚪生长发育和变态的影响
施林强,毛 飞,马小梅*
(杭州师范大学生命与环境科学学院,浙江 杭州 310036)
实验用泽陆蛙蝌蚪为模型动物检验食物水平充足的环境中密度对其生长、变态和存活率的影响.密度水平设置为30只/L、60只/L和120只/L.研究发现,蝌蚪的体长、尾长和体重的增长速率随密度的增大而减小;高密度显著降低早期的存活率,但随后的存活率不受影响;高密度显著降低蝌蚪的变态率,表明高密度抑制了蝌蚪的发育从而延长变态时间;变态体重不受密度影响.该研究推测高密度环境降低蝌蚪生长速率是导致其变态率降低的主要原因,变态体重不受密度影响可能是实验动物在水体稳定且食物充足的生境中对高密度导致的变态延迟代价的一种补偿;泽陆蛙蝌蚪生长发育的适宜密度环境可能需低于30只/L.
泽陆蛙; 蝌蚪; 密度;发育; 变态
0 引 言
两栖类的变态特征(如变态大小、变态时间等)会影响其变态后的适合度.变态时间短且变态个体大的幼体往往具有较高的适合度,表现为在陆地环境中,比变态时间长且变态个体小的幼蛙生长速率大、达到性成熟时间短、性成熟个体大以及存活率高等优势[1-4].变态时间缩短能延长幼蛙在第一个冬季来临前的活动时间以便获得充足的食物,加快生长发育速率,较早地达到性成熟的个体大小或顺利完成越冬,实现自身利益的最大化;相反,变态时间长且变态个体小的幼体由于活动季节的缩短,食物资源的竞争加剧,导致存活率下降.
种内竞争是影响两栖类变态特征的一个重要因子,大量的实验室和自然条件下的研究发现,高密度培养环境下的个体变态时间长、变态个体小、生长速率和存活率低[5-9].然而,一些模型认为种内竞争加剧时,生长速率的降低能诱导变态[10-11].在一定的密度范围内,适宜的密度大小能加速蝌蚪的变态,过低或过高的密度均能延长变态时间[12].此外,Relyea[13]还研究发现,林蛙(Ranasylvatica)蝌蚪在较高种内竞争环境中形态发生适应性改变,增强了竞争能力.
该研究利用泽陆蛙(Fejervaryalimnocharis)蝌蚪为实验材料,在充分饱和给食的条件下,检验密度对蝌蚪生长、发育和变态特征的影响,探究变态特征的最佳密度水平.研究变量包括蝌蚪体长、尾长和体重的生长速率、存活率、变态率和变态大小(体重).
1 材料与方法
1.1 泽陆蛙成体的采集与繁殖
于2009年4月30日在浙江富阳市郊农田(30°01’N,119°45’E)捕获约200只泽陆蛙成体,第二天运回杭州师范大学两栖爬行动物实验室.经测量获得形态数据后将它们投放入室外3个蛙池(长 × 宽 × 高 = 3.6 × 1.8 × 0.8 m3)中饲养.池中间放置露出水面的水泥预制板和若干石棉瓦片以供其栖息和躲藏,保持池中水深约12 cm.在饲养期间,每天18:00用面包虫或蚯蚓饱和喂食.
在7月上旬进入繁殖高峰期,每晚观察泽陆蛙的抱对情况,及时收集所产的卵放于盛2 L曝气水的塑料盆中并置于25 ℃的恒温室内进行孵化培养.光照周期为12 L:12 D,每天06:00自动开启.该研究共采用4窝同一天所产的卵进行密度实验.
1.2 蝌蚪的密度实验
待约3 d蝌蚪出膜后(Stage 25,Gonser,1960),每窝随机选择10只大小基本一致的蝌蚪测量其体长(SVL,snout-vent length)、尾长(TL,tail length)和所有个体的总重并计算平均体重(MBM,mean body mass).每窝随机捞取一定数量的蝌蚪进行密度实验.密度设置如下:每个塑料盆(直径 × 高 = 26 cm × 9 cm)中分别放入10(30只/L)、20(60只/L)和40(120只/L)只蝌蚪.盆中水深约1.3 cm,每个密度重复2次.每天09:00用鱼饲料饱和给食,并保持不同密度下的投饵量基本相同,每天18:00用预温过的曝气自来水换水一次.实验在25 ℃恒温室内进行,光照周期为12 L:12 D,每天06:00自动开启.
蝌蚪饲养过程中,每隔12 d记录每盆中蝌蚪的成活数,并随机捞取10只蝌蚪(<10只的全部测量)用游标卡尺和电子天平分别测量它们的SVL、TL和总重.当蝌蚪进入变态期,每天保持多次观察并及时取出已进入变态的蝌蚪(Stage 42,前肢伸出)放入小塑料盆.待其尾巴完全消失后,测量幼蛙的体重.待少量蝌蚪陆续进入变态,于第60 d停止形态指标的测量,继续饲养直到第93 d实验结束.
1.3 数据分析
该研究中,SVL、TL和MBM的生长速率 =(第60 d的测定值-第1 d的测定值)/60(d).所有数据均用Kolmogorov-Smirnov与Bartlett分别检验正态性与方差均质性.数据分析用单因子方差分析(one-way ANOVA),Tukey’s检验进行多重比较.所有数据分析用Statistica 5.0统计软件包完成,描述性统计值用平均值 ± 标准误(范围)表示,显著性水平设置为α = 0.05.
2 结 果
2.1 密度对蝌蚪生长的影响
泽陆蛙卵在25 ℃的恒温室内3 d后发育历期达到Stage 25,此时蝌蚪(N= 40)的体长为2.9 ± 0.04 mm,尾长为5.12 ± 0.07 mm,体重为0.006 ± 0.01 g.
蝌蚪在不同密度培养下均呈现出先快后慢的生长特点.在培养前12 d SVL和TL迅速增长,随后生长速率逐渐减小,直至进入变态期.密度对SVL和TL生长的影响显著,在每个生长测定点,不同密度下的SVL和TL差异显著(allP< 0.0001),均表现为SVL和TL随培养密度的增大而显著减小(图1a和图1 b).密度显著影响蝌蚪的体重增长,且随着生长高低密度间的体重差异日益扩大(图1 c).在每个生长测定点,蝌蚪体重随密度的增大而减小,表现为30只/L培养的体重最大,120 只/L培养的体重最小,60只/L培养的体重处于两者之间,30只/L与120只/L培养的蝌蚪体重差异均显著(图1 c).
培养60 d内,蝌蚪SVL、TL和MBM的生长速率随密度的增大而显著减小,且任意两密度间的差异均显著(allP< 0.0001)(图2).
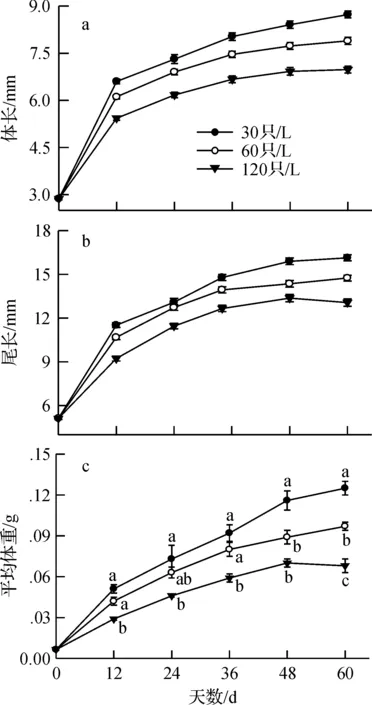
图1 泽陆蛙蝌蚪在不同密度培养下体长、尾长和平均体重随时间的变化Fig.1 Changes in snout-vent length,tail length and mean body mass with culture time at different densities of F.limnocharis tadpoles
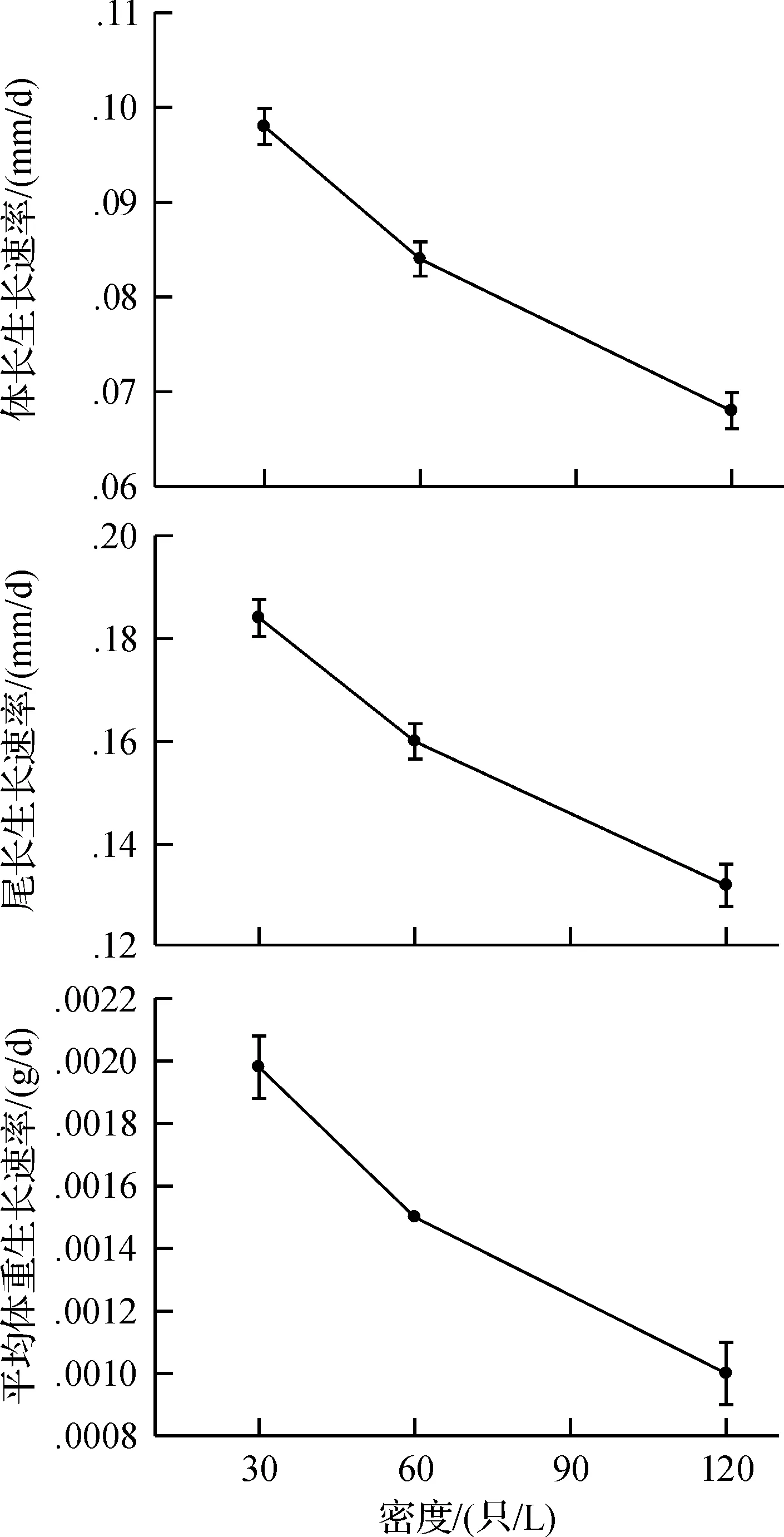
图2 密度对泽陆蛙蝌蚪生长速率的影响Fig.2 The effects of density on growth rates of snout-vent length,tail length and mean body mass in F.limnocharis tadpoles
2.2 密度对蝌蚪存活率的影响
密度对蝌蚪早期(前12 d培养期间)的存活率具有显著的影响(F2,20= 3.68,P= 0.043),随密度的增大存活率显著降低,在培养的第12 d,30只/L、60只/L和120只/L培养的存活率分别下降了10%、19.4%和25%.培养12 d后,密度对存活率的影响变小,表现为每个生长测定点的存活率差异均不显著(allP> 0.05),但低密度培养下的存活率始终大于高密度培养下的存活率(图3),至培养第60 d,30只/L、60只/L和120只/L培养下的存活率分别为79%、73%和65%.
2.3 密度对蝌蚪变态特征的影响
密度显著影响蝌蚪的变态率,30只/L培养下的蝌蚪存活率显著大于60只/L和120只/L培养下的存活率,60只/L和120只/L培养下的存活率无显著差异;在93 d的培养时间内,30只/L、60只/L和120只/L培养下蝌蚪的变态率分别为40%、5.8%和0.8% d(表1).至第93 d实验结束时,30只/L、60只/L和120只/L培养下的变态个数分别为24、7和2只.密度对变态幼蛙的体重无显著影响(表1).
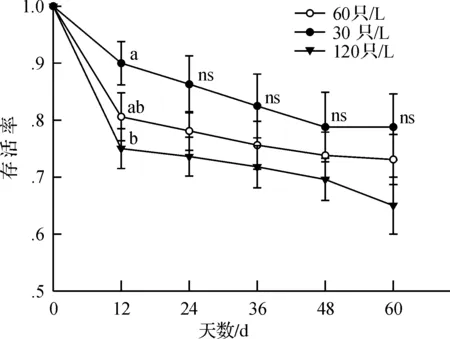
图3 泽陆蛙蝌蚪在不同密度培养下存活率随时间的变化Fig.3 Changes in survival rate with culture time at different densities of F.limnocharis tadpoles
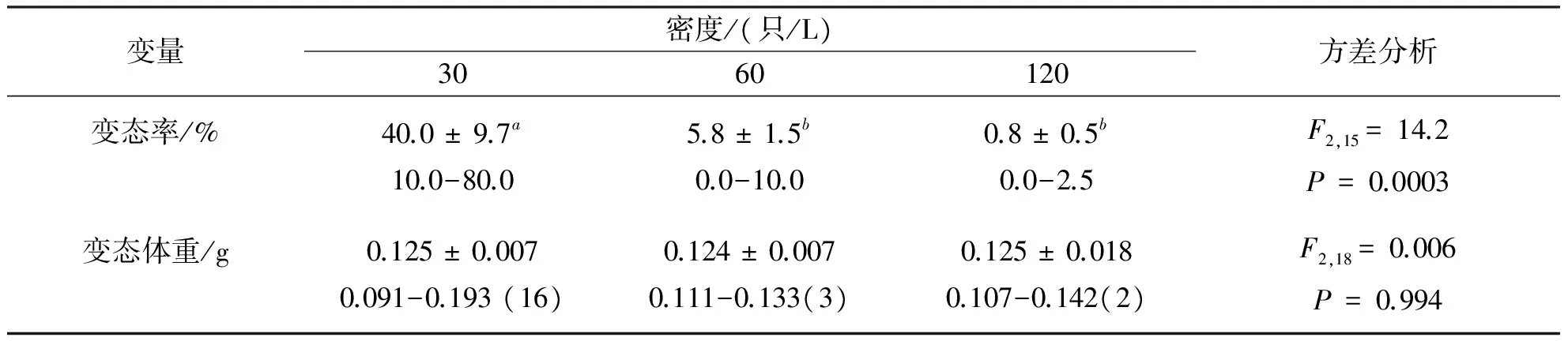
表1 泽陆蛙变态特征的描述性统计值Table 1 Descriptive statistics of metamorphic traits in F.limnocharis tadpoles
不同上标小写字母表差异显著(Tukey’s post hoc test,α=0.05,a>b)
3 讨 论
该研究发现,在饱和给食的条件下,高密度培养降低泽陆蛙蝌蚪的生长速率,却不影响蝌蚪的存活率和变态幼蛙的体重.蝌蚪的变态率随密度的增大而显著降低可间接地推测高密度培养环境降低了蝌蚪的发育速率,延长了变态时间.上述结果与一些相关的研究结果一致[12,14],然而,该研究得出密度不影响存活率和变态个体的体重这一特征与大多数的研究结果不同,即其他的研究发现,高密度培养的蝌蚪比低密度培养时存活率低、变态个体小,体重轻[5,8,9,15-17].
密度对蝌蚪变态时间的影响依赖于生长速率.高密度培养环境引起种内竞争加剧,生长速率下降,尽管在食物资源丰富的条件下[7,18],造成蝌蚪发育速率减慢,变态时间延长.研究发现,当控制不同密度培养下楔头蛙(Ranasphenocephala)蝌蚪的生长速率一致时,高密度培养能加速蝌蚪变态.该研究中,泽陆蛙蝌蚪在高密度培养下生长速率显著降低(图2),可能是造成变态时间延长以及变态率低的原因.Rot-Nikcevic等[7]发现林蛙蝌蚪能利用视觉评价种内竞争强度的变化,在食物资源不受限制的条件下,高密度培养导致生长和发育速率的降低,而Rot-Nikcevic等[19]发现林蛙蝌蚪受到的种内竞争加剧由视觉和触觉同时施加时,其生长速率减慢,但发育速率显著增大,这说明林蛙蝌蚪同时利用视觉和触觉信息来调整其发育速率,响应环境变化,且这种应激模式受到肾上腺酮激素(corticosterone)的控制[20];然而,不同的物种利用这些机制的程度存在较大的区别.因此,该实验动物的食物资源尽管不是制约因子,但仍表现出高密度培养下的生长速率降低,这也可能是蝌蚪通过视觉和(或)触觉对高密度培养时种内竞争加剧的一种评估,从而响应环境变化的结果.此外,高密度培养下的强干扰竞争(interference competition)(如大个体蝌蚪释放的化学物质、摄食取代)也可能是种内竞争加剧、生长速率减小的原因之一[21].
密度对变态大小的影响往往表现为变态个体体长或体重随密度的增加而减小[4,6,8,16,22].该研究实验动物变态大小不受密度的影响,可能是该物种对自然条件下食物资源丰富和积水期较长水体的一种适应;由于高密度环境下的个体生长速率缓慢,延长变态时间而获得与低密度环境下变态个体一样的大小,可能是对变态延迟代价的一种补偿.
该研究中,泽陆蛙蝌蚪在60 d的生长期间,第12 d已表现出高密度环境对生长速率的抑制,且在随后的生长过程中,密度间的形态差异日益显著,表明高密度环境对早期生长速率的负作用持续影响后期的生长.然而,不同的是,密度显著影响第12d的存活率,而随后的存活率在密度间无差异,这可能是发育早期的蝌蚪比生长后期的蝌蚪易受环境干扰,且高密度环境下较高的死亡率导致种内竞争减弱的结果.然而,泽陆蛙蝌蚪在最低密度(30只/L)培养下的存活率仅为79%,表明低于30只/L的密度可能更有利该物种的生存.
4 结 论
该研究表明,在食物资源充足且水体稳定的生境中,泽陆蛙蝌蚪的生长和发育速率随密度的增大而减小;高密度环境降低蝌蚪的变态率,但变态大小不受密度影响,可能是高密度环境下生长速率减小造成的;高密度对蝌蚪生长的影响在早期(第12 d)已显现,且随生长日趋明显;高密度显著降低早期的存活率,但随后的存活率不受影响;泽陆蛙蝌蚪更适宜生存的密度环境可能需低于30只/L.
[1] Smith D C.Adult recruitment in chorus frogs:Effects of size and date at metamorphosis[J].Ecology,1987,68(2):344-350.
[2] Morey S,Reznick D.Effects of larval density on postmetamorphic spadefoot toads (Speahammondii) [J].Ecology,2001,82(2):510-522.
[3] Altwegg R,Reyer H U.Patterns of natural selection on size at metamorphosis in water frogs[J].Evolution,2003,57(4):872-882.
[4] Relyea R A,Hoverman J T.The impact of larval predators and competitors on the morphology and fitness of juvenile treefrogs[J].Oecologia,2003,134(4):596-604.
[5] Scott D E.Effects of larval density inAmbystomaOpacum:An experiment large-scale field enclosures[J].Ecology,1990,71(1):296-306.
[6] Loman J.Density regulation in tadpoles ofRanatemporaria:A full pond field experiment[J].Ecology,2004,85(6):1611-1618.
[7] Rot-Nikcevic I,Taylor C N,Wassersug R J.The role of images of conspecifics as visual cues in the development and behavior of larval anurans[J].Behavioral Ecology and Sociobiology,2006,60(1):19-25.
[8] Belden L K,Rubbo M J,Wingfield J C,etal.Searching for the physiological mechanism of density dependence:Does corticosterone regulate tadpole responses to density?[J].Physiological and Biochemical Zoology,2007,80(4):444-451.
[9] Provenzano S E,Boone M D.Effects of density on metamorphosis of bullfrogs in a single season[J].Journal of Herpetology,2009,43(1):49-54.
[10] Wilbur H M,Collins J P.Ecological aspects of amphibian metamorphosis[J].Science,1973,182(4119):1305-1314.
[11] Day T,Rowe L.Developmental thresholds and the evolution of reaction norms for age and size at life-history transitions[J].American Naturalist,2002,159(4):338-350.
[12] Browne R K,Pomering M,Hamer A J.High density effects on the growth,development and survival ofLitoriaaureatadpoles[J].Aquaculture,2003,215(1-4):109-121.
[13] Relyea R A.Competitor-induced plasticity in tadpoles:Consequences,cues,and connections to predator-induced plasticity[J].Ecological Monographs,2002,72(4):523-540.
[14] Hailey A,Sookoo N,Hernandez E,etal.The influence of density and ration level on culturedPhysalaemuspustulosustadpoles,and the mitigation of a crowding effect by soil substrate[J].Applied Herpetology,2007,4(3):261-277.
[15] Martinez I P,Alvarez R,Herraez M P.Growth and metamorphosis ofRanaperezilarvae in culture:Effects of larval density[J].Aquaculture,1996,142(3-4):163-170.
[16] Girish S,Saidapur S K.Density-dependent growth and metamorphosis in the larval bronze frogRanatemporalisis influenced by genetic relatedness of the cohort[J].Journal of Biosciences,2003,28(4):489-496.
[17] Indermaur L,Schmidt B R,Tockner K,etal.Spatial variation in abiotic and biotic factors in a floodplain determine anuran body size and growth rate at metamorphosis[J].Oecologia,2010,163(3):637-649.
[18] Flores-Nava A,Vera-Munoz P.Growth,metamorphosis and feeding behaviour ofRanacatesbeianaShaw1802 tadpoles at different rearing densities[J].Aquaculture Research,1999,30(5):341-347.
[19] Rot-Nikcevic I,Denver R J,Wassersug R J.The influence of visual and tactile stimulation on growth and metamorphosis in anuran larvae[J].Functional Ecology,2005,19(6):1008-1016.
[20] Glennemeier K A,Denver R J.Role for corticoids in mediating the response ofRanapipienstadpoles to intraspecific competition[J].Journal of Experimental Zoology,2002,292(1):32-40.
[21] Steinwascher K.Interference and exploitation competition among tadpoles ofRanautricularia[J].Ecology,1978,59(5):1039-1046.
[22] Stamper C E,Stevens D J,Downie J R,etal.The effects of competition on pre- and post-metamorphic phenotypes in the common frog[J].Herpetological Journal,2008,18(4):187-195.
TheEffectsofDensityonGrowth,DevelopmentandMetamorphosisofFejervaryaLimnocharisTadpoles
SHI lin-qiang,MAO Fei,MA Xiao-mei
(Hangzhou Key Laboratory for Animal Adaptation and Evolution,Hangzhou Normal University,Hangzhou 310036,China)
The experiment used tadpoles ofFejervaryalimnocharisas model to test the effects of density on their growth,metamorphosis and survival rates when the food sufficient.The density regime was set at 30,60 and 120 individuals per liter water.These results show that the increasing rates of snout-vent length,tail length and body mass of tadpoles decrease with increased density.In early time of this experiment,the survival rate decrease significantly under high density,whereas not affects by density thereafter.The percentage of metamorphosed tadpoles decline with high density which suggests that high density restrains tadpoles’ development and prolongs the time for metamorphosis.The metamorphosis mass is not affected by density.The paper speculated that the decrease of growth rate caused by high density was the main reason for low percentage of metamorphosed tadpoles.The fact that metamorphosis mass was not affected by density was possibly as a compensation for the costs of prolonged metamorphosis time under high density when tadpoles had stable water and sufficient food.The most suitable density for growth and metamorphosis of tadpoles possibly would be lower than 30 individuals·L-1.
Fejervaryalimnocharis; tadpole; density; development; metamorphosis
2010-01-12
浙江省自然科学基金项目(Y307435).
施林强(1984—),男,浙江富阳人,生态学专业硕士研究生,主要从事动物的进化生态学研究.
*通信作者:马小梅(1962—),女,浙江杭州人,高级实验师,主要从事动物学实验和科研工作.E-mail:mxm@hznu.edu.cn
10.3969/j.issn.1674-232X.2011.05.009
Q958.1
A
1674-232X(2011)05-0429-06
